
2
Existing Understanding of the Environmental Context for Hominin Evolution
Before presenting prioritized research initiatives designed to increase understanding of the interaction between human evolution and Earth’s environmental history, it is important to assess what is known today about the fundamental steps in hominin evolution, the history of climate and other environmental parameters in areas where hominins evolved, the contribution of other biota recovered with hominins to understanding climatic impacts, and the history of human modifications of ecosystems. In each of these sections, the focus is on Africa from the time of the earliest hominins through to their first dispersal out of Africa at about 1.8 Ma. Descriptions of the youngest part of the record extend beyond Africa. These descriptions are necessarily brief and represent the committee’s understanding of existing broad scientific consensus at the time of publication of this report. Although there is ongoing scientific research in each of these disciplinary areas to address the myriad scientific uncertainties and inconsistencies that will always exist in predominantly data-limited fields, the overview summaries presented here will not attempt to analyze or present the details of these uncertainties and inconsistencies.
MAJOR EVENTS IN HUMAN EVOLUTION
The pattern and process of human evolution can be described on the basis of a combination of comparative anatomy, the fossil record, and primate and human genetics (Kimbel and Martin, 1993). Comparative anatomy, even in Darwin’s time, indicated a close relationship between humans and African apes (Huxley, 1863), and this has been confirmed by comparative genetic analyses. Although the branching order of the ape family tree—gibbons, orangutans, gorillas, chim-
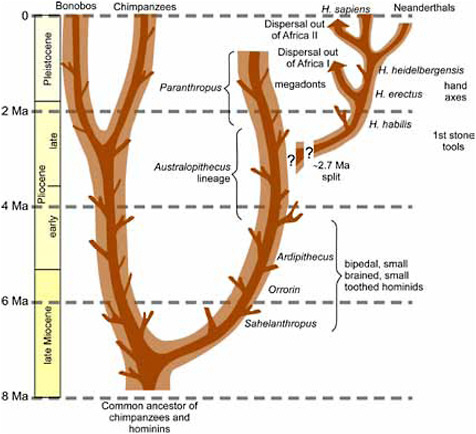
FIGURE 2.1 Highly simplified summary of hominin evolution over the past 8 Ma—the numerous terminating “twigs” schematically illustrate evolutionary “dead-ends.”
panzees, humans—is firmly established, the dates of these branching splits are less certain (Kumar et al., 2005). The earliest fossils of the human lineage, after the split from the common ancestor of the chimpanzees (Figure 2.1), are fragmentary and the dates of some remain imprecise. A distorted cranium from Chad, Sahelanthropus tchadensis, has a reduced snout compared with apes, and skull characteristics that are sometimes taken to indicate bipedality (Brunet et al., 2002; Zollikofer et al., 2005). The site from which this specimen comes (Koro Toro on Figure 2.2) has recently been dated to between 6.8 and 7.2 Ma (Lebatard et al., 2008), and this estimate is consistent with the faunal evidence. Other early fossils from Kenya (Orrorin tugenensis; Senut et al., 2001) consist of fragmentary jaws and limb bones with dates of 5.7 to 6.0 Ma. Although there is debate about the exact relationship between O. tugenensis and later hominins, recent anatomical analyses of the skeleton (Richmond and Jungers, 2008) indicate that this species was habitually bipedal—a uniquely hominin trait. So by 6 Ma, our earliest
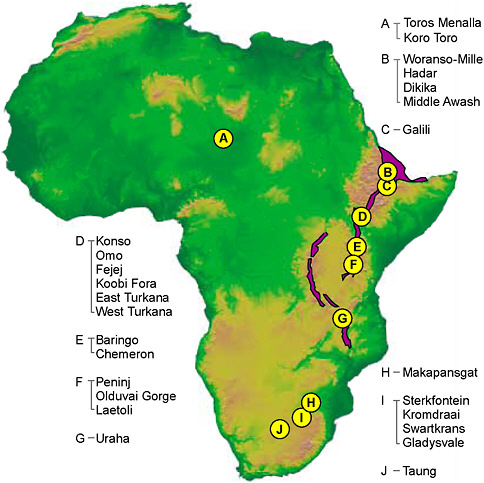
FIGURE 2.2 Geographic distribution of major exploration sites for hominins older than 1.8 Ma (i.e., prior to the first dispersal of H. erectus out of Africa). East Africa Rift System shown in purple. SOURCE: Digital elevation model image courtesy National Oceanic and Atmospheric Administration National Geophysical Data Center.
ancestors had split from the chimpanzee lineage and become adapted to bipedal locomotion, which is the major difference that separates us from great apes.
The fossil record of hominins between 6 and 3 Ma is patchy, but samples from Ethiopia, Kenya, Tanzania, and Chad record several bipedal hominins that have been placed in the genera Ardipithecus and Australopithecus. The early part of this hominin record (5.8 to 4.4 Ma) is represented by Ardipithecus, which had acquired some features seen in later Australopithecus, but which still exhibited primitive traits seen in African great apes (White et al., 1994, 1995, 2009 and associated articles).
The species in the genus Australopithecus all have larger molar and premolar teeth and thicker enamel than their predecessors (Ward et al., 1999, 2001;
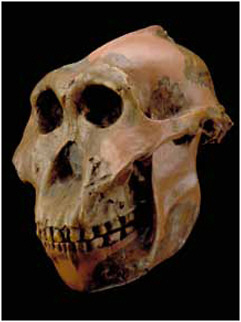
FIGURE 2.3 Replica of a 1.8-Ma Paranthropus boisei cranium found by Mary Leakey in 1959 at Olduvai Gorge, shown with a replica of a 1.2-Ma mandible of the same species from Peninj, Tanzania. SOURCE: Image courtesy Human Origins Program, Smithsonian Institution.
White et al., 2006), and gradual change from A. anamensis to A. afarensis has been documented (Kimbel et al., 2006). Although their food processing anatomy differed, this lineage of bipedal hominins had brain to body mass ratios that are about the same as those of extant great apes. Their limb proportions differed from those of both chimpanzees and humans, and their pelvic and hip structure suggests a somewhat different mode of bipedal locomotion from that of our own genus Homo. Confirmation of bipedal locomotion comes from fossilized footprints at Laetoli in Tanzania (Leakey and Hay, 1979). Australopithecus species exhibited differences in body size and canine dimensions between males and females (i.e., sexual dimorphism). A. afarensis is well known from cranial and postcranial parts and includes the famous partial skeleton ”Lucy” from ~3.2 Ma. A related hominin—A. africanus—is well known but poorly dated from South African cave sites. One individual is of a nearly complete skeleton (Clarke, 1998; 2002), which promises to deliver much important information about this southern form. The youngest member of this genus, A. garhi, was recovered from 2.5-Ma deposits in Ethiopia (Asfaw et al., 1999), but little is known about it except that the trend throughout this lineage to larger jaws and teeth continued.
Extremely large-toothed hominins appear in the record around 2.7 Ma. These are sometimes placed in Australopithecus but are more commonly assigned to their own genus, Paranthropus (Figure 2.3). These “robust” creatures, so-called
because of their massively sized jaws and teeth, may be direct lineal descendents of A. afarensis. The earliest species from East Africa is P. aethiopicus, known only from a single cranium and other isolated skeletal parts. This species evolved into P. boisei, and fossils of this younger species are relatively common in East Africa (Constantino and Wood, 2007). A similar species, P. robustus, is found in cave sites in South Africa; these fossils are also found with bone fragments that were used to dig both tubers and termites (Backwell and D’Errico, 2001; Pickering et al., 2004). Although we have only discovered limited numbers of skeletal bones of these robust-jawed hominins, they seem to have been very similar to the earlier Australopithecus in their postcranial adaptations (the skeletal features aside from the skull, jaw, and teeth). It is likely that the larger jaws and teeth were used for chewing very hard foodstuffs. Paranthropus appears to have become extinct at about 1.2 Ma or shortly after, at the same time that several other African mammal species became extinct. Although there are many characteristics and capabilities that remain unknown, one thing is clear; Paranthropus existed at the same time and in the same areas as the earliest members of our own genus, Homo. Their co-occurrence is the firmest evidence for different species of hominins existing together.
Behavioral evidence for the existence of our own genus, in the form of stone tools, predates any Homo fossils so far discovered, and although stone tools are commonly associated with the genus Homo throughout its existence it is not possible to be completely certain that Paranthropus did not make all or some of the earliest tools. Stone tools referred to as Oldowan technology,1 as old as 2.6 Ma, have been found in Ethiopia (Semaw et al., 2003). The earliest definite Homo fossil is a 2.3-Ma maxilla (upper jaw) from an Oldowan archaeological site at Hadar, Ethiopia (Kimbel et al., 1996), which shows a steeper facial profile and a broader palate than Australopithecus species. By about 2.0 Ma, fossils of early Homo and sites with animal bones and stone tools are relatively common. However, it is important to emphasize that although these hominins have been assigned to the genus Homo, this does not imply that they were very much like modern humans in anatomy and behavior. Consequently, we should be wary of attributing any particular human behavior or physiology to early Homo without strong evidence. We do not know which particular Australopithecus species gave
rise to Homo, although there have been suggestions that A. garhi was the precursor species (Asfaw et al., 1999).
The remains of Homo habilis are known from East Africa (Tobias, 1991; Wood, 1991), and possibly also from South Africa (Grine et al., 1993); this species is either very variable in cranial capacity and palate shape, or there are two species present. In general, this species has a mixture of primitive features as well as some that foreshadow those of the later H. erectus. Sexual dimorphism in body size was strong, like that of the preceding Australopithecus species.
The appearance of early Homo erectus at about 1.9 Ma is marked by changes in the limb skeleton that make it nearly indistinguishable from that of modern humans; these changes have been associated with the capacity for long-distance running (Bramble and Lieberman, 2004). This species is the first hominin to disperse out of Africa. Dispersal of H. erectus to present-day Georgia, where it is found at Dmanisi, apparently took place shortly after the first evidence for its existence in East Africa (Gabunia et al., 2001; Antón and Swisher, 2004), and evidence of the dispersal of Homo to East Asia by about 1.8 Ma is documented in China and Indonesia (Antón and Swisher, 2004; Zhu et al., 2004, 2008). This dispersal out of Africa is widely believed to have been facilitated by a major behavioral shift to increased hunting and meat consumption (Shipman and Walker, 1989). These hominins were quite variable in their cranial capacity (Spoor et al., 2007), probably due to sexual dimorphism. Studies of enamel formation show that their life history was like that of African apes rather than humans—they grew up quickly and died young (Dean et al., 2001). It is interesting to note that the Acheulean stone tool culture that is thought to typify H. erectus, which included the handaxe, had not been developed by the time their first fossils occurred. This species apparently used Oldowan technology until the Acheulean was invented at about 1.6 Ma; their dispersal to Eurasia, for example, took place without handaxes. The earliest strong evidence for the use of controlled fire occurs about 800 ka (Goren-Inbar et al., 2004).
Homo sapiens—in the form of skulls and skeletons that are practically indistinguishable, even in brain size, from those of modern people—appears first in Africa about 200 ka (McDougall et al., 2005). Before this event there are many fossils that are usually allocated to H. heidelbergensis, as well as other more arcane names. Some of these fossils from Europe appear to be the ancestors of the Neanderthals, a group of hominins that evolved in the glacial climates of Eurasia. Other fossils from Africa are most likely the ancestors of Homo sapiens (White et al., 2003), and others recovered in Asia may not have had any descendants.
Many behaviors that are usually attributed to modern people have left traces in the archaeological record of Africa from about 200 to 100 ka. These innovations appeared in the following order: (1) shellfishing; (2) fine stone blades, grindstones, and ochre use; (3) stone points; (4) long-distance exchange of material; (5) fishing, bone tools, barbed points, mining, and etched items to
record information; (6) microlithic blades and bead ornaments; and (7) images (McBrearty and Brooks, 2000; Marean et al., 2007).
Modern humans emerged in Africa long before the Neanderthals became extinct in Europe. A dispersal of H. sapiens out of Africa occurred at about 60 ka, with modern humans reaching as far as Australia at that time. Humans arrived in the Americas only recently, at about 30-15 ka (Meltzer, 2003). The reasons for this late arrival are still unclear, but certainly during the most recent glacial maximum the climatic conditions were severe in eastern Siberia, the Bering Strait region, and the western portion of arctic North America.
In summary, the major evolutionary events in human evolution are:
-
The split from chimpanzees at 8-6 Ma.
-
The development of bipedal locomotion, probably occurring at the split.
-
The slow evolutionary change to bigger teeth, thicker enamel, and reduction of canines that characterize a 5-Ma-long lineage from Sahelanthropus and Orrorin (if those are not the same as Ardipithecus), through Ardipithecus, to Australopithecus, and finally to Paranthropus.
-
A splitting event between 3 and 2.5 Ma that produced Homo from an Australopithecus ancestor.
-
The development of stone tool technology at about 2.6 Ma.
-
The origin of a more carnivorous species, Homo erectus, at about 1.9 Ma.
-
The first dispersal by hominins out of Africa, by 1.8 Ma.
-
The development of the Acheulean stone tool culture at about 1.6 Ma.
-
An increase in cranial capacity in H. heidelbergensis at about 500 ka.
-
The origin of Homo sapiens at about 200 ka.
-
The origin of symbolic language.
-
The successive innovations in culture and lifestyle that led to the second dispersal event out of Africa at about 60 ka.
-
Expression of symbolic language in cave paintings and sculptures by about 60-30 ka.
-
The domestication of plants and animals within the last tens of thousands of years in different parts of the world.
-
The ever-accelerating spread and dominance of humans over global ecosystems in the last few thousand years.
There are, of course, many problems of interpretation and several major disagreements about human evolution that cannot be explored in this brief summary. One of these disagreements is that several researchers have followed the view of Stephen Jay Gould that human evolution must have produced a “bushy” evolutionary tree (Gould, 1994), whereas others prefer a simpler tree (White, 2003). Despite a large body of literature documenting extensive intraspecific variation in higher order primate morphology, there are still disagreements among researchers concerning the expected range of variation within fossil species.
MAJOR EVENTS IN EARTH SYSTEM HISTORY ASSOCIATED WITH HUMAN EVOLUTION
The environmental context for hominin evolution—the environmental characteristics that prevailed in the areas where hominins evolved—fundamentally reflects the interplay between Earth’s orbital parameters, tectonism, and the biogeochemical processes that controlled greenhouse gas concentrations. The tectonic elements set the stage by delineating land and ocean, including oceanic gateways and land bridges, as well as controlling regional and local topography. Earth’s orbital variations determined the amount of solar radiation any location on the Earth’s surface received at a given time and season. Greenhouse gas concentrations controlled the very large scale characteristics of planetary temperature and regional moisture balance, and the state of the cryosphere. Together, these produced the interaction of atmospheric air masses with topographic and oceanic effects that controlled the specific regional climates that impinged on evolving hominins.
The history of East African tectonics and orography is dominated by the development of the East African Rift System (Tiercelin and Lezzar, 2002) (see Figure 2.2). Prior to inception of the East African Rift System, most of northeastern Africa was a low-lying landscape of deeply weathered terrain. Major eruptions of flood basalt and rhyolites created the Ethiopian Plateau around 30 Ma (Wolfenden et al., 2005), marking the onset of rifting along the East African Rift System. By 10 Ma, active rifting had propagated southward for 4,000 km, from the Gulf of Suez to the Mozambique Channel. Rifting in the Turkana basin, the Red Sea, and the Gulf of Aden were all coincident with the onset of Afar volcanism. The rift system has two distinct branches, an older and more volcanically active eastern branch, active since the Oligocene and occupied today by many small alkaline/saline lakes, and a younger (late Miocene) western branch, which has experienced much less extensive volcanic activity and is occupied by large and mostly freshwater lakes. Unlike the eastern branch of the Rift Valley, where southward propagation of rifting is well documented, the western branch appears to have developed along its entire length at about the same time between 8 and 12 Ma. Both rift branches are surrounded by major uplifted mountain ranges, which have acted since the late Miocene to intercept moisture and create rainshadows along their leeward (western) flanks (Sepulchre et al., 2006). This rift valley system and the flanking mountains regulated the extent of aridity in various parts of tropical Africa, as well as the occurrence of water resources upon which early hominins would have relied.
The development of the Nile River system was an additional significant element in the history of hominin evolution on the continent. There is no evidence for a Nile drainage system prior to the early Miocene, when major fluvial deposits began to accumulate south of the present Nile delta (Said, 1993). Today the Nile is fed by both the Blue Nile that drains the Ethiopian Highlands and the White
Nile that drains Lake Victoria and the lakes occupying the northern reaches of the western rift. The Ethiopian Highlands would have been the sole significant drainage area in the early stages of Nile development before rifting began in the western arm of the rift valley at around 15 Ma.
The other major element of African tectonic history for hominin evolution was the gradual and complex collision of Africa with Eurasia that began at about 15 Ma (Burke, 1996). This event had several critical implications for hominins. First, it generated land connections between Africa and Eurasia at various times that provided biogeographic corridors for terrestrial organisms, including hominins, to move between the two continents. And second, when coupled with variations in global sea level, this collision caused the episodic closure of the Straits of Gibraltar and isolation of the Mediterranean from the Atlantic during the late Miocene. This resulted in the evaporation of the Mediterranean Sea (the “Messinian salinity crisis” of about 7-5 Ma; see Rouchy et al., 2006), profound regional climatic changes, biogeographic corridors between Africa and Eurasia, and changing conditions for ecosystems in the region (e.g., van der Made et al., 2006). The sea-level history of the Mediterranean also strongly influenced the history of the Nile, a major biogeographic corridor between tropical Africa, the rift system, and Eurasia.
Regional and Global Controls on African Climate Change
An understanding of how African climate has varied through time and may have influenced human evolution requires a broad understanding of how the African climate system works today. The climate of tropical Africa is impacted primarily by three air masses—the northeasterly and southeasterly trade wind systems, which penetrate East Africa from the Indian Ocean and converge on the Intertropical Convergence Zone (ITCZ), and the Atlantic-derived westerly African monsoon, which extends into the African interior where it converges with the easterly trade winds along the Congo Air Boundary. This boundary is positioned along the western rift for much of the year, where orographic effects promote enhanced rainfall. The pattern of rainfall throughout East Africa is complex. In general the seasonal rain belt migrates north and south with the ITCZ, modified by regional orography, vegetation, and the energy exchange between extratropical and tropical regions (Leroux, 2001). Rainfall throughout most of the East African rift valley is derived from moisture off the Indian Ocean, and is strongly influenced by regional differences in sea surface temperatures (Hastenrath et al., 1993). South Africa is affected by the westerly wind belt as well as by tropical climate. The interannual variability of rainfall in South Africa is out of phase with that in tropical East Africa, and tends to be relatively dry in El Niño years (Goddard and Graham, 1999).
Climate variability during the Neogene is expressed on various timescales, each of which may have been important for hominin evolution. Over the longest
timescales, climate trends over millions of years—the global cooling trend and the growth of polar ice sheets—set the stage for the overall evolution of hominins. On shorter timescales, Earth orbital (Milankovitch) processes were critical for controlling aridification cycles in Africa, which could have influenced hominin distribution, adaptation, and local water resource availability. And at millennial and shorter timescales, abrupt climate events could have influenced the demography of individual hominin populations, local extinctions, and population distribution around water resources. Each of these timescales is considered below.
Late Neogene Aridity Trends in Africa
Terrestrial and marine paleoclimate records have been interpreted to show that subtropical African climate has, over the past 2-3 Ma, progressively become more arid with an expansion of grasslands. This longer term aridity trend appears to have been superimposed on the higher frequency precessional wet-dry cycles, although the precise nature of this superposition remains unclear. Marine sediment records of African climate changes have provided several independent lines of evidence for this latest Neogene aridification trend. Depositional fluxes of eolian sediment off West and East Africa reveal increases coincident with the 2.8 Ma onset of Northern Hemisphere glaciation, and the dust cycles in these records exhibit changes in variability at 2.8 Ma, 1.7 Ma, and ~1 Ma coincident with similar changes in high-latitude ice volume variability (Tiedemann et al., 1994; deMenocal, 2004). A recent reanalysis of the timing and nature of these transitions suggests that there may be significant differences in the timing of these transitions between western and eastern North African sites (Trauth et al., 2009). Several studies have shown a close correspondence between the pacing of high-latitude glacial cycles and African climate using dust fluxes (Clemens et al., 1996; Tiedemann et al., 1994), dust grain size (Matthewson et al., 1995), pollen records (Dupont and Leroy, 1995), and biomarker tracers of tropical African rainfall (Tierney et al., 2008). Moreover, the pacing of the dust cycles is in phase and coherent with the oxygen isotopic record of glacial/interglacial climate changes, with three- to fivefold increases in dust fluxes during glacial maxima at sites off northeast and northwest Africa. Marine pollen records similarly document expansion of vegetation adapted to drier conditions during glacial maxima, and a general aridity trend after the mid-Pliocene (Dupont and Leroy, 1995).
Although terrestrial paleoclimate data have the potential to provide critical ground truth information on long-term climate trends near the fossil localities, continuous paleoclimate records are rare from East African terrestrial sequences because of active faulting, erosion, and nondeposition. The sparse existing records broadly support the view that East African climate changed from warmer, wetter conditions in the late Miocene and early Pliocene to a more seasonally-contrasted, cooler, and drier (and perhaps more variable) climate during the late Pliocene (after ~3 Ma). Pollen from fossil sites in northeastern Africa indicates
shifts to cooler and drier vegetation types after ~2.5 Ma. Stable isotopic analyses of pedogenic carbonates from the Turkana and Olduvai basins indicate progressive replacement of closed forest woodland by open savannah grasslands between 3 and 2 Ma, with further increases after 1.8 Ma, 1.2 Ma, and 0.6 Ma (Cerling and Hay, 1988; Cerling, 1992; Levin et al., 2004; Wynn, 2004) (Figure 2.4). However, sufficient C4 grass biomass was present to support a diverse community of grazers from ~7 Ma to the present in many key hominin-bearing localities (Cerling et al., 1999, 2005; Levin et al., 2008).
A compelling new line of evidence for increasing African aridity is provided by stable isotopic analyses of plant wax biomarker compounds preserved in marine sediments off equatorial and northeast Africa. The Gulf of Aden is the closest marine basin to hominin fossil localities in northeast Africa, and low-resolution plant wax biomarker analysis from ocean cores recovered from the Gulf of Aden document a clear trend toward more open C4 vegetation, commencing between 3 and 2 Ma (Feakins et al., 2005) (Figure 2.4). Together, the terrestrial and marine records demonstrate that African savannah grasslands became an increasingly prominent component of the landscape after the mid-late Pliocene.
Orbital Precession Forcing of African Climate
The geological record provides abundant evidence of orbital precession (see Box 2.1) having been the persistent pacemaker of African climate changes. In the recent geological past, semiarid parts of North Africa have experienced periodic and dramatic changes in moisture availability due to regulation by orbital precession of African monsoonal circulation. During the early Holocene (North) African Humid Period, the modern Saharan Desert was nearly entirely vegetated (Jolly et al., 1998), and the landscape was dotted with numerous large and small permanent lakes supporting abundant large mammalian fauna (Roberts, 1998). Collectively, terrestrial and marine paleoclimate records present compelling evidence that African climate periodically alternated between wetter and drier conditions throughout the late Neogene.
The varying concentrations of eolian dust exported from regional North African source areas to offshore northwest and northeast Africa, noted above, have also recorded these precessional wet-dry cycles (Tiedemann et al., 1994; Clemens et al., 1996; Larrasoaña et al., 2003; deMenocal, 2004), and sedimentary records from East Africa lake deposits show extreme climatic variability with rapid shifts between wet and dry conditions (Trauth et al., 2007). Terrestrial paleolake deposits in Ethiopia, Kenya, and Tanzania contain a record of the precessional wet phases, but the preserved record of African humidity cycles is far less complete (Ashley, 2007; Kingston et al., 2007).
The climate history south of the equator in tropical East Africa also reflects a strong precessional cycle that is out of phase with North Africa. The Lake Malawi basin, at the southern end of the East African Rift Valley, was relatively
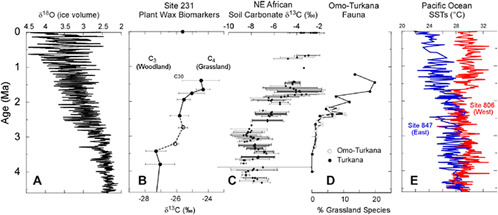
FIGURE 2.4 Long-term African climate trends (B-D) shown with oceanic temperature and ice volume records (A, E). (A) Marine δ18O record showing the onset and amplification of high-latitude glacial cycles after 3 Ma (Lisiecki and Raymo, 2005); (B) Carbon isotopic composition of Deep Sea Drilling Program Site 231 terrestrial plant wax biomarkers (Feakins et al., 2005); (C) East African soil carbonate δ13C data (Cerling, 1992; Cerling et al., 1994; Wynn, 2004); (D) Turkana Basin faunal shifts documenting C4 vegetation (grassland) expansion after the mid-Pliocene (Bobe and Behrensmeyer, 2004); and (E) Pliocene-Pleistocene evolution of equatorial Pacific Ocean sea surface temperature (SST) gradients documenting the absence of the modern-day east-west SST gradient during the middle and late Pliocene (Wara et al., 2005), an oceanographic boundary condition consistent with more humid conditions in East Africa (Cane and Molnar, 2001; Brierley et al., 2009).
|
BOX 2.1 Earth’s Orbital Variation and Climate Variability—Milankovitch Climate Cycles and the Mediterranean Sapropel Record In the early 20th century, Milutin Milanković, a Serbian scientist, demonstrated that the amount and distribution of solar energy reaching the Earth is predictably and cyclically related to variations in the Earth’s orbit. Three types of orbit variation are particularly important for controlling the amount and seasonality of solar energy reaching different parts of the Earth—precession cycles, controlled by changes in the orientation of the earth’s axis; obliquity cycles, reflecting the degree of tilt of the Earth’s axis; and eccentricity cycles, the degree to which the Earth’s orbit around the sun differs from a circle. For over 30 years, paleoclimatologists have recognized the importance of changes in—and interactions between—these various cycles as regulators of the Earth’s climate. This is because the distribution of solar radiation reaching the Earth’s surface not only controls how warm a given region will be at a given time of year, but also how atmospheric circulation redistributes moisture across the globe. The Mediterranean sapropel record provides some of the most compelling evidence that North African climate has responded to orbital precession regulation of monsoonal climate since at least the late Miocene (Hilgen, 1991; Hilgen et al., 1995; Lourens et al., 1996; Kroon et al., 1998). When orbital precession invigorated monsoonal rainfall, the Nile drainage basin captured and directed runoff to the eastern Mediterranean where the excess supply of freshwater stratified Mediterranean surface waters (Sachs and Repeta, 1999). This stratification inhibited deep water convection and the deep Mediterranean became anoxic during African humid events, which are marked by intermittent (~5-ky duration) periods of organic-rich sediment accumulation in the deep basins of the central and eastern Mediterranean (Rossignol-Strick et al., 1982). The periodic regular |
dry during the early Holocene North African Humid Period (Finney and Johnson, 1991; Castaneda et al., 2007). Drill cores from Lake Malawi reveal a 150,000-year history of climate that differs considerably from that of the Congo Basin, with droughts occurring on a precessional frequency prior to 75 ka (Cohen et al., 2007; Scholz et al., 2007).
Millennial Scale and Shorter Climate Variability in Africa
African climate records show substantial variability on interannual to millennial timescales, and climate events on these scales may have had an important effect on hominin evolution by influencing water resources available to individual populations or through the occurrence of catastrophic events. During the Holocene period, when Greenland ice cores indicate a generally stable high-latitude climate, lakes located from the Afar region of Ethiopia to tropical West Africa experienced dramatic rises and falls in lake level, reflecting the recurrence of
|
ity of these sapropel layers (Figure 2.5) reflect Earth’s eccentricity and precession with remarkable fidelity throughout the late Neogene (Hilgen, 1991), and this periodicity, after correlation with rapidly-deposited beds containing hominin fossils, has been used as the basis for accurately dating the fossils (McDougall et al., 2008). Precession-paced changes in ocean salinity estimated from isotopic analyses of sediment cores near the mouths of major North African rivers (e.g., Nile, Niger, and Congo) have also been used to document the timing and duration of these African humid periods (Kroon et al., 1998; Rohling, 1999; Weijers et al., 2007; Weldeab et al., 2007). 
FIGURE 2.5 Left panel shows an Ocean Drilling Program core from Site 967 in the eastern Mediterranean Sea, dramatically illustrating cyclicity between sapropel layers (dark) interspersed with carbonate ooze (light). The right panel shows, on an outcrop scale, approximately 50 sapropel layers of late Miocene age (~9 Ma) from the Gibliscemi section in Sicily. SOURCE: Core photograph from Emeis, Robertson, and Richter et al. (1996). Outcrop photograph courtesy Dr. Frits Hilgen, University of Utrecht, Netherlands. |
severe, persistent drought alternating with floods (Talbot and Delibrias, 1980; Street-Perrott and Perrott, 1990; Gasse, 2000).
Human population effects resulting from climate events on these timescales are well documented from the Holocene (e.g., Kropelin et al., 2008). Millennial-scale events have been little explored for older parts of the records, although a long core from Lake Tanganyika (Tierney et al., 2008) and a drill core from northern Lake Malawi (Brown et al., 2007) both provide evidence for abrupt and dramatic climatic shifts on this timescale during MIS-3.
PALEOBIOLOGICAL CONTEXT OF HUMAN EVOLUTION
An important contribution to understanding how human evolution might have been affected by climate and/or habitat change is provided by the other biota that existed both with hominins and apart from them. As climate altered vegetation habitats (determined by rainfall, evapotranspiration, soils, and other aspects of the earth system), this habitat modification applied selective pressures
|
BOX 2.2 Environmental Indicator Fauna Associated with Hominin Fossils Paleoenvironmental interpretations of hominin habitats have become increasingly more sophisticated over time; whereas early paleoanthropologists typically inferred habitats based on one or two species, it is now more common to use fossil groups or lineages (e.g., Vrba, 1995) or the entire faunal assemblage (e.g., Bobe and Eck, 2001; Reed, 2008) recovered with hominins as the basis for habitat inference. The following mammal groups are often found at hominin localities; taken individually, they provide rather variable indications of the environmental attributes of their surroundings, but they become much more precise environmental indicators when considered in totality (e.g., see Figure 2.6). Bovids, i.e., Antelopes, are probably one of the best indicators of habitat because of their particular adaptations to different kinds of savanna vegetation. Different bovid species are adapted to specific environments, and have been ubiquitous 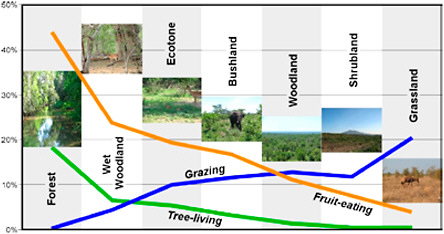
FIGURE 2.6 Variation in the ratio of adaptations (food types and habitats) in modern mammal communities for different ecosystems in game reserves and parks in Africa. SOURCE: Courtesy of Kaye E. Reed. |
on the fauna that used these habitats, leading to new adaptations, speciation, or extinction. The adaptations that are evident in the faunal assemblages recovered with hominins provide a valuable line of evidence that can be used to reconstruct paleoenvironments and track environmental change.
One of the best ways to reconstruct the habitat and ecosystems in which hominins lived is to analyze the fossil flora associated with hominin sites—the pollen, leaf fossils, and other plant parts. However, this direct evidence of vegetation is typically better preserved only at younger paleoanthropological sites, but for older paleoanthropological localities—where fossil plants are less common—the fossil fauna often must be used to interpret habitat because faunal specimens are ubiquitous and nearly always associated with hominin fossils.
|
in Africa since the end of the Miocene. For example, the bovids that are related to modern wildebeests usually indicate open, seasonal, grassy environments. Cervids are mammals, including deer, reindeer, and moose, where the males have antlers that are shed yearly and females usually have no headgear. These animals, found mostly in the Northern Hemisphere, are adapted to various habitats. Reindeer are usually inferred to indicate a tundra habitat. Suids are the pigs of Africa, Europe, and Asia. Early animals in this group were omnivores, but they underwent an adaptive radiation during the middle Pliocene and many species became adapted to grazing. The teeth of the omnivores and grazing suids are distinctively different, allowing specific habitats to be inferred. Giraffids are fairly common, usually long-necked browsing species that are typically associated with African savannas. Although there are only two non-overlapping species today, as many as four giraffid species coexisted in the same area in the Plio-Pleistocene. Different neck lengths of giraffids provide habitat information by indicating adaptations to different tree heights. Hippopotamids are large, water-dwelling ungulates that indicate the presence of rivers and lakes in the tropical African Plio-Pleistocene. Occasionally they have been recovered from European sites (e.g., London) during warm interglacial periods, indicating that tropical conditions extended as far north as the British Isles. Equids (horses) are mammals that possess a single toe today, but some past lineages had three toes. Equids originated in the Americas, but dispersed to Eurasia and Africa in the Miocene. Two groups are known from Africa—hipparions, which have been recovered from most Plio-Pleistocene hominin localities; and Equus, which migrated into Africa from Eurasia at about 2.5 Ma. The entire group of fossil species are grazers that existed in fairly open grasslands, but whether these represent wet or dry grasslands is debated. Rhinocerotids are mammals with three toes and one or two nasal horns. Two rhino species are found at almost all African hominin localities, and Eurasian rhino species have been recovered with Neanderthals and modern humans in Europe and with modern humans in North Africa. Both African rhino species can be found in the same types of savanna habitats, although the Black Rhino generally prefers more bush and tree cover. |
Floral Evidence Tree height and spatial distribution strongly affect animal species distributions and richness today, and it is reasonable to assume that they did so in the past. The spatial distribution spectrum ranges from closed habitats, with high tree densities, to open habitats with more dispersed trees. Variability of tree density is one of the terrestrial manifestations of climate change, and habitat change has been interpreted as an important selective force influencing many of the events in human evolution.
Faunal Evidence There are a number of ways to understand and interpret faunal information from the past. In some cases, individual fossil species can provide information on the habitat from which they were recovered through an understanding of how a skeleton moved, or by determining an animal’s diet
from morphological examination or isotope examination of the teeth. However, broader-based faunal community analyses using fossil fauna assemblages (see Box 2.2) can often provide a more reliable indication of the habitats of hominin fossil sites (Vrba, 1995; Reed, 1997, 1998, 2008), as well as an understanding of species turnover resulting from climate change (Behrensmeyer et al., 1997; Bobe and Eck, 2001; Alemseged, 2003).
Faunal assemblage changes—the shifts in species or faunal types through time or across space—indicate changes in vegetation structure. Species abundance or presence/absence data can also be used to understand faunal evolution associated with climate change; for example, the increased length of elephant teeth through time indicates development of more open, arid habitats with dust-coated leaves. The total assemblage of fossils, coupled with our understanding of adaptations in individual fossil species, can be used to interpret how habitats change through time (Reed, 1997, 2008). Finally, presence/absence patterns of fauna across the globe can provide evidence for dispersal patterns, and an understanding of when movement within and among continents was possible. This biogeographic information is a key element for understanding hominin movement within Africa and dispersal beyond Africa.
Vegetation and Faunal Changes Through Time in Africa
Savanna environments, where grasses are the dominant ground cover (White, 1983), can range from closed woodlands to open grasslands. Although temperature and rainfall are the major determinants of savanna occurrence elsewhere around the globe, African savannas and their vegetation types respond primarily to the amount and seasonal distribution of rainfall, with soil type as a less important influence. The seasonal distribution of rainfall is so important that seasons are described as either rainy or nonrainy, rather than the spring, fall, summer, and winter of higher latitudes. It is reasonable to infer that rainfall imposed a similar control on savanna distribution in the African past, leading to the conclusion that the forests from which middle Miocene apes have been recovered reflect high rainfall rates with the rainfall distributed evenly throughout the year. Cerling et al. (1997) and Brunet et al. (2002) suggested that the earliest hominins lived in a mixed grass and tree habitat or a more open lake margin setting. These reconstructions are based on soil isotopes and plant remains, as well as on the fauna recovered with the hominins. Mammalian and other fauna associated with Orrorin (Senut et al., 2001) and Ardipithecus (WoldeGabriel et al., 1994, 2001) fossils indicate that these hominins probably existed in predominantly closed woodlands, or even forests.
There were long-term changes in the seasonal distribution of rain in the later Miocene, with leaf analysis showing that the length of dry seasons increased by many months in the middle to late Miocene, and then shortened again near the Mio-Pliocene boundary (Jacobs and Deino, 1996; Jacobs et al., 1999). This
climate change was mirrored by a change in faunal species compositions from a variety of sites in East Africa, with the extinction of many middle Miocene taxa and the appearance of modern lineages of mammalian taxa in the middle to late Miocene, at ~8.5–6.5 Ma (Hill, 1995). At the same time, plant isotopic records from the Tugen Hills, Kenya (6.8 Ma) indicate that some of the fauna, which were adapted to more open habitats, were consuming grasses that grow in a much cooler environment than is present in Africa today. Consequently, there appear to have been heterogeneous landscapes during the late Miocene of East Africa, including forests, woodlands, and grasslands. Fossil vertebrates show minor changes through time, from those adapted to living in forests to those adapted to more open country. For example, elephant and equid species changed from browsing (eating leaves and bushes) to grazing (eating grasses) during this time period.
Some of the earliest hominins from the Mio-Pliocene boundary period may have inhabited forests and/or closed woodlands (WoldeGabriel et al., 1994; Haile-Selassie, 2001; Senut et al., 2001), although Sahelanthropus at ~7 Ma was recovered with fauna that indicate a wooded grassland environment close to a lake (Brunet et al., 2002). Many fossil mammal assemblages in Africa show evidence of faunal interchange with Eurasia. The fauna associated with Ardipithecus at Adu-Asa, in the Middle Awash region of northern Ethiopia, are more closely associated with African, rather than Eurasian fauna, although there were some immigrant Eurasian taxa recovered. Interestingly, the other African site with fauna most similar to Adu-Asa is at Langebaanweg, on the west coast of South Africa. Langebaanweg also contains the Eurasian fauna and is approximately half a million years younger than Adu-Asa. Although no hominins have been recovered from Langebaanweg, the site is important for understanding climatic and biogeographic patterns during this important time period. This is one of the few periods in which there is evidence for Eurasian immigrant faunas and for shared mammals between eastern and southern Africa. Similarly, the fauna recovered from Chad with Sahelanthropus has African affinities, but seems to also have several endemic species, a trend that continued into the middle Pliocene.
Faunal evidence, based on large numbers of colobine monkeys, has been interpreted to indicate that Ardipithecus ramidus (4.4 Ma) from the Middle Awash region of Ethiopia lived in somewhat closed habitats (White et al., 2006), whereas isotopic evidence for this species (5.2-3.9 Ma) from the Gona region further to the north indicates a predominance of grass-eating animals (Levin et al., 2008). Together, the evidence may indicate that A. ramidus lived in environments that varied over time or favored areas that consistently contained both riparian forest and nearby open habitats. Soil isotope data indicate that Australopithecus anamensis lived in relatively open habitats in the Turkana Basin, Kenya (Wynn, 2000).
Faunal reconstructions of habitats of middle Pliocene localities bearing A. afarensis, A. africanus, A. bahrelghazali, and the fossils attributed to Kenyanthropus platyops (3.6 to ~2.8 Ma), indicate varied environments ranging from closed
to open woodlands and bushlands, as well as wooded grasslands (Andrews, 1989; Harris, 1991; Reed, 1997; Leakey et al., 2001; Harris et al., 2003). Reconstructions show that K. platyops lived in wet and somewhat closed habitats (Leakey et al., 2001), and there are different habitats and faunal assemblages through time that are associated with the A. afarensis sites at Hadar in Ethiopia (Bonnefille et al., 2004; Reed, 2008) and Laetoli in Tanzania (Andrews and Bamford, 2008). The mammals from Hadar show a shift from dominantly browsers to mammals that lived in more arid environs and ate grasses and shrubs. In addition, gastropods from Hadar indicate that the length of the dry season increased across the latter part of A. afarensis existence (Hailemichael, 2000).
The australopithecine species from the Koro Toro site in Chad is associated with more open, lake-margin habitats, with fauna that have both East and North African affinities as well as endemic taxa (Geraads et al., 2001). Fauna recovered from the cave sites of Makapansgat and Sterkfontein in South Africa indicate two disparate habitats; Makapansgat, the older locality (~3.0 Ma), has fauna representing heterogeneous habitats that include floodplain, woodland, bushland, and forest. Sterkfontein, on the other hand, contains fauna that indicate very open grassland habitats, as well as plant species that indicate forest. Irrespective of the habitats associated with this southern hominin taxon, the faunal distribution during this time period suggest that habitat or climatic mechanisms contributed to the biogeographic isolation of South Africa from eastern Africa—the fauna from Makapansgat and Sterkfontein are more similar to each other, although separated by 500 ky, than to fauna recovered from sites of similar ages in East Africa.
Therefore, early to middle Pliocene habitats were heterogeneous, but with a tendency toward more open habitats broadly similar to modern African savannas. The adaptations evident in the fossil faunas show a great deal of variability with respect to habitat type, but there are more arboreal, and thus forest-associated, animals found in the late Miocene than the middle Pliocene. Through the early to middle Pliocene there is also evidence for environmental events that apparently caused species isolation not only in distant parts of the continent, but also between some of the East African sites.
In all sites for which there are data, the time period surrounding ~2.8 Ma shows some faunal turnover, but the rate and pattern of the faunal changes are different at each site (Bobe and Eck, 2001; Alemseged, 2003; Reed, 2008). There have been suggestions that these species turnovers were pan-African in nature, and caused by a global climate changes (Vrba, 1988, 1995), although the fossilrich Turkana Basin exhibits a different pattern and timing of species turnover (Behrensmeyer et al., 1997) than would be expected with this hypothesis. By about 2.3 Ma, there was an almost complete replacement of mammalian taxa at many localities (e.g., Alemseged, 2003; Reed, 2008). New hominin species (e.g., earliest Homo species) also appeared at this time. However, the lack of continuity of faunal and floral records makes habitat interpretations more difficult and/or less reliable—there are fauna associated with each new hominin species
recovered, but sites with continuous faunal records have discontinuous records of hominins. However, there was an overall trend in the fauna from ~2.8 to 1.8 Ma throughout Africa indicating a shift from more closed to more open habitats. At 1.8 Ma, not only were these faunas in Africa replaced by many grazing taxa, but also hominins had dispersed out of Africa for the first time. However, the stable isotope record of mammalian diets does not show a perceptible change in diets over this interval for most mammal groups (Cerling et al., 2005).
African fauna from 1.8 to ~1.0 Ma in hominin-bearing localities indicates fairly open, grassland habitats, and this interpretation is supported by paleosol isotope data (Cerling, 1992). In general, the percentages of grass and trees in African landscapes fluctuated during this time, but never reverted to closed conditions. South African cave sites from which Paranthropus robustus has been recovered indicate open woodland environments with some floodplains (Reed, 1997). There is some indication that the preservation of P. robustus fossils tended to occur only during relatively arid intervals, when sediments and bones more easily entered subterranean caves, and it has been proposed that southern Africa, in general, represented a more stable landscape than those associated with East African hominins (de Ruiter et al., 2009). P. boisei habitats in East Africa have been interpreted as wetter and more wooded (Shipman and Harris, 1988), and Homo erectus is associated with woodland and grassland habitats (Reed, 1997). Woodlands expanded to replace some of the grasslands at about 1.6 Ma, but there is another grassland peak at ~1.0 Ma (Cerling, 1992).
Vegetation Structure and Faunal Changes through Time Outside Africa (post-1.8 Ma)
After hominins dispersed from Africa towards the north, they interacted with different types of habitats and fauna. Some of these habitats were in more temperate climates with pronounced temperature seasonality, whereas other habitats, such as those in Southeast Asia, have completely different faunal assemblages compared with those in Africa, despite similarly tropical conditions.
There are few long continental records for Eurasia for this post-1.8-Ma period. Marine sediment records from the North Atlantic show the fundamental orbital-cycle pacing of glacial/interglacial cycles and long-period trends for this interval, as recorded in the marine oxygen isotope record (Raymo, 1994). By inference, this record provides a broad context for the timing of glacial advances and retreats in both Eurasia and North America. The most recent interglacial commenced around 125 ka, with the onset of glaciation at ~115 ka, glacial maximum at ~20 ka, and rapid initial retreat at ~15 ka. There are extensive land records for this period, and they provide a basis for inferring the large changes in climate—and related changes in flora and fauna—that influenced all of Eurasia during this period. Climate model simulations of global climates at glacial maxima and
interglacial maxima also help bound the range of spatial and temporal climate variability over this vast area and time.
The earliest evidence for dispersal of hominins out of Africa is at ~1.8 Ma, when Homo erectus appears in the Caucasus Mountains of Georgia, and perhaps slightly later in mainland China and on Java, Indonesia. In western Asia, the Homo erectus site of Dmanisi has few African species, but many typical Eurasian species (Gabunia et al., 2000). The fauna from Dmanisi indicates an environment consisting of forested river margins with open steppe between rivers, and the lack of many African-derived species (Tappen et al., 2007) suggests the dispersal of H. erectus from Africa was unusual for the time. Although the Java site is wet tropical today, the fauna possibly indicate a drier woodland environment at 1.8 Ma (Storm, 2001). In general, orangutans and gibbons, characteristic of Southeast Asian rain forests, are not present in faunas from the Homo erectus sites of Trinil and Ngandong on Java. Fossil pollen and animal remains from the 1.7-million-year-old site of Yuanmou, South China, indicate that Homo was associated with a diverse habitat of open vegetation, bushland, and forest (Zhu et al., 2008), while Nihewan Basin archaeological sites dated at 1.66 to 1.32 Ma indicate an environment that varied between open grassland and more dense vegetation (Zhu et al, 2004). Homo erectus is known until at least 250 ka in China, and therefore must have adapted to the more extreme warm-cool climate variations in that area.
Little is known about the many habitats associated with H. heidelbergensis. H. neanderthalensis is associated with cold climates and cold-adapted faunas such as reindeer, bison, cave bears, and cave lions. Neanderthals dispersed into the Middle East at several times during their existence, and the associated fauna provides some interesting insights. First, the cold-adapted fauna are not found there, indicating that Neanderthals also dispersed without the other fauna. Second, the fauna recovered at the Neanderthal localities in the Middle East are the same as the fauna recovered with Homo sapiens from the same area, although there are indications that Neanderthals focused on exploiting fewer species, whereas H. sapiens used a broader range of animals as food (Reed and Fish, 2005). Early H. sapiens (~200-125 ka) in Africa were associated with fauna that indicate either open grassland (grazing bovids and equids) or fynbos2/Mediterranean habitats (small browsing bovids) (Rector and Reed, 2009). With the expansion of Homo sapiens out of Africa at ~60 ka, it is evident that they were no longer restricted or limited by any particular habitat or climate. Although modern faunas are essentially in place from about 200 ka in all parts of the globe, many large animals disappear during the late Pleistocene with their extinction representing some combination of habitat change and hunting.
HUMAN MODIFICATION OF ECOSYSTEMS
The record of human modification of the environment spans at least 2.5 million years. During this period, changing climates would have resulted in changes and uncertainties in the availability of critical resources (e.g., food and water). Hominins that survived such uncertainty must have adapted to these challenges, with key evolutionary innovations altering the ways in which human ancestors interacted with their surroundings. New technologies variously involving the use of stone, the intensified hunting of animals and reaping of wild plants, and the potential to build shelters, to clear landscapes using fire, and to play a role in extinctions of other organisms, together with the developing ability to communicate and plan coordinated activity—all set the stage for a fundamental change in human ecology involving the transition from mobile hunting-gathering to food production and the emergence of human-dominated ecosystems. Framed in the context of late Pliocene and Pleistocene climate change, the capacity to make tools, exploit new foods, control fire, build durable shelters, and organize complex social activity reflect evolutionary responses that enabled human ancestors to survive and adapt to environmental risks and uncertainties (Potts, 1996b, 1998). The initially simple capacities to alter their immediate surroundings proved so successful that they enabled humans to spread over the planet and thus, ultimately, to induce environmental change on a global scale.
The most profound human modifications of ecosystems resulted from the transition from food foraging (e.g., hunting-and-gathering) to food production (e.g., farming, herding). This fundamental change in how humans acquired food involved a transition from dependence on wild food sources sought and found each day, involving much of the population, to dependence on food that could be grown and stored by a much smaller subset of the population (Flannery, 1986; Zeder, 2006). The investment in fields and food production meant that originally highly mobile groups became sedentary. Populations grew in size due to the existence of a more stable food supply, which also enabled some members of the population to become specialized craftsmen, artists, inventors, religious and political figures, along with the many other roles that people adopt in modern society (Diamond, 1997). The following is a summary of several distinct developments over the long course of human evolution that provided the antecedents to this critical transition.
Technology At present, the oldest documented stone tools attributable to hominins are dated to nearly 2.6 Ma (Semaw et al., 2003). Pliocene toolmaking involved the manufacture of sharp-edged stones (by using other stones as percussors) and the use of rocks for crushing and pounding. Even the simplest cutting and crushing activities enabled early hominin toolmakers to gain access to new higher-quality foods (e.g., animal fat and protein, or buried tubers and roots) (Potts, 1996b). From this simple technological beginning, a large array of food
resources became available for exploitation by the relatively small-brained ancestors of living humans.
Similar stone toolmaking techniques were also practiced by the first populations of the genus Homo to disperse from Africa to Eurasia, by about 1.8 Ma (Rightmire et al., 2006; Zhu et al., 2008; Potts and Teague, in press). This basic technology persisted for hundreds of thousands of years, with few innovations. The innovation of striking large flakes and the emergence of Acheulean handaxes, by about 1.6 Ma, does not seem initially to have changed the way early hominins interacted with their surroundings, and the environmental factors that may have influenced the temporally and spatially patchy record of handaxe-focused technology are not well understood. Handaxes do, however, become very well crafted by at least 500 ka, and were useful in a wide range of tasks (Schick and Toth, 1993). The oldest known thrusting spears, made of wood, are from ~400 ka, indicating that human ancestors could reliably hunt animals by at least that time. Innovations in stone technology began to occur at a slightly faster rate after 300 ka, as smaller, more diverse, and easily transported tool kits began to replace handaxes and other large cutting tools that had dominated stone technology for the previous 1.3 million years. Early populations of Homo sapiens had developed the capacity to invent specialized tools (e.g., projectile points by at least 105 ka, and bone harpoon points by ~80 ka) that were useful in capturing dangerous and fast-moving prey.
Concentrations of Refuse and Intensification of Human Activities The human tendency to collect wastes is evident in the simple beginnings of the archaeological record. Early archaeological sites consist of concentrations of toolmaking refuse (stone chips) and food remains (typically butchered and broken animal bones). A mobile, foraging lifestyle meant that early toolmakers could follow resources as habitats changed seasonally in the short term, or more dramatically over longer time spans. This mobile existence also meant that hominins could move away from concentrations of wastes as they accumulated.
As modern humans began to develop new technologies, especially after 100 ka, some groups began to exploit and manage animal herds that were predictable in their behavior and to intensify their use of wild plants, particular cereal grains. Human populations were able to displace unwanted carnivores and other animals. In some places, this intensification of human activity entailed settling into an area where resources could be managed or cultivated; this less mobile way of life occurred thousands of years prior to the emergence of food production. Ultimately, the development of agriculture led to a certain degree of control over the food supply and an investment in the landscape that gradually led human populations to abandon hunter-gatherer mobility in favor of settlement. Building upon the human capacity to concentrate waste, these developments inevitably led to the buildup of wastes, pollutants, etc., that characterize human-modified ecosystems in the present.
Fire and Shelters The control of fire and the building of durable shelters were critical means by which mid-Pleistocene hunter-gatherers altered their immediate environment and this also occurred long before the emergence of agriculture. The oldest definitive hearths have been dated at approximately 790 ka (Goren-Inbar et al., 2004), and shelters that were sufficiently durable to be preserved in the archaeological record date from approximately 400 ka (deLumley, 1969; Schick and Toth, 1993). The construction of hearths and shelters enabled hominins to modify the temperature of their immediate surroundings, alter the digestive properties of cooked food, and to distance the group from harsh conditions beyond the shelter. These developments coincided with a period of heightened amplitude of glacial/interglacial oscillations, and thus may reflect the ways in which altering the immediate environment proved beneficial physiologically to individuals and socially to groups (Potts, 1996b). Ultimately, the use of fire also led humans, mainly after 100 ka, to modify entire landscapes as a means of hunting or land clearance to promote new plant growth (e.g., Lentz, 2000; Miller et al., 2005).
Sophisticated Symbolic and Cognitive Behavior The evolution of complex mental capabilities and language had a strong impact on how our species interacted with its surroundings. These developments provided an adaptive advantage in Homo sapiens by enabling social groups to trade resources over long distances and to cope with variations in food, water, and other critical resources in the face of climate change. These cognitive capabilities are indicated by early symbolic artifacts, such as pigments used for coloring, simple etching of objects, and the presence of decorative shell beads. These types of objects indicate an ability to code information symbolically—the essence of language. These artifacts first occur in the African archeological record between 285 ka and 70 ka, associated with the early evolution of Homo sapiens (Barham, 2002; Henshilwood et al., 2002; d’Errico et al., 2005). By approximately 130 ka, artifacts made from rocks from at least 300 km away suggest that human social networks were sufficiently complex to engage in long-distance exchange of high-quality stone and other resources (McBrearty and Brooks, 2000). By at least this date, therefore, our species manifested complex mental behavior and highly coordinated social activity. These developments laid an important part of the foundation for large-scale human impacts such as agriculture, trade, and cities, which have profoundly altered the relationship between humans and natural environments.
SUMMARY ENVIRONMENT-EVOLUTION CHRONOLOGY
The major features of human evolution and the major features of Earth’s climatic evolution over the past 8 million years can be integrated to form a chronological summary, summarized in Table 2.1. This provides the context for the recommendations for the future research and outreach activities that are presented in subsequent chapters.
TABLE 2.1 Summary of Critical Intervals in Earth System and Hominin Evolutionary History for Africa During the Late Neogene.
|
Interval |
Climate Events |
Sea-Level Events |
Tectonic Events |
Hominin Evolutionary Events |
Archaeological Record Events |
Fossil Record Events (other than hominins) |
|
8-4 Ma |
|
|
|
|
|
|
|
4-2 Ma |
|
|
|
|
|
|
|
Interval |
Climate Events |
Sea-Level Events |
Tectonic Events |
Hominin Evolutionary Events |
Archaeological Record Events |
Fossil Record Events (other than hominins) |
|
2-0.5 Ma |
|
|
|
|
|
|
|
0.5-0.0 Ma |
|
|
|
|
|
|




























