
4
Emerging Infections, Tick Biology, and Host–Vector Interactions
Today ticks inhabit almost every continent, with the number of species worldwide topping 850. Ticks have proven resilient and persistent in the environment, and the fossil records suggest that they originated 65–146 million years ago (see Olsen and Patz; Paddock and Telford, Appendix A).
The recognized number of important diseases transmitted by ticks has been growing over the past 30 years (see Paddock and Telford, Appendix A). The emergence and increased incidence of several major tick-borne diseases (TBDs) has been attributed to specific human activities and behaviors that disrupt ecosytems (see Paddock and Telford, Appendix A). Increases in human population and demographic shifts have brought dramatic changes in the distribution and composition of natural habitats, as people modify the land to create living spaces for agriculture or for recreation (see Munderloh and Kurtii, Appendix A). These changes mean that people and animals interact at many more interfaces, creating new opportunities for the transmission of zoonotic diseases, including TBDs. For example, habitat fragmentation can alter the movement of hosts that carry TBDs, the dynamics of disease transmission, and biodiversity (see Appendix A). Global environmental changes and other abiotic and biotic factors also help shape the ecology of TBDs and their emergence and reemergence.
Fortunately, new molecular tools and analytical techniques such as gene sequencing and analysis have enabled scientists to gain insights into tick biology and have resulted in a better understanding of TBDs. New technologies have also revealed a diverse microbial community associated with ticks that include viruses, bacteria, protozoans, and fungi. These microbes may act as symbionts (interacting closely, often to the benefit of the tick),
pathogens, and transient commensals (colonizing the tick without marked detrimental effects), or as pathogens (see Clay and Fuqua, Appendix A).
In this chapter, five scientists examined the natural history of ticks and their wildlife and domestic hosts; outlined the contributions of animal health experts to understanding human TBD; explored genetic diversity among pathogens, vectors, and hosts; and showed how scientists investigate the microbial community found within the ticks to better understand the human risk for tick-borne diseases.
EMERGING AND REEMERGING TICK-BORNE INFECTIONS: GENETIC MANIPULATION OF INTRACELLULAR TICK-BORNE PATHOGENS
Ulrike G. Munderloh, D.V.M., Ph.D. Department of Entomology, University of Minnesota
Ticks are efficient vectors of multiple pathogens due to their potential interactions with several different vertebrate hosts during their life cycle. As a result, they have the opportunity to acquire a large array of different types of organisms that are present in the blood of these hosts. The microbial community in ticks includes viruses, bacteria, protozoa, and fungi, and serve as symbionts, commensals, and pathogens. In fact, the organisms that comprise the tick microbiome vastly outnumber recognized human pathogens. This microbial community can influence the acquisition, transmission, and virulence of human pathogens. Furthermore, as the tick feeds for extended periods, it interacts with its vertebrate host and has the ability to suppress the host’s immune system by dampening down the immune response and binding up antibodies that the host might have made in an attempt to rid itself of the blood-sucking parasite. These attributes ensure that a pathogen can be acquired from or transmitted to a bite site that is suppressed and immunologically inactive.
Anaplasma phagocytophilum shares a vector, the black-legged tick, with Lyme disease spirochetes, a vector that is expanding its range, which helps to explain the increasing incidence of human granulocytic anaplasmosis. The white blood cells, specifically the neutrophils, are infected in reservoir mammalian hosts in the peripheral blood, and in lungs, heart, spleen, and gut. Animals also serve as models to account for the multiple signs of disease that infected people may present. There is a need to understand how these pathogens can survive and flourish in a broad range of mammalian hosts and a number of organs within the host, as well as in vector ticks. This can be done with new techniques to analyze how microbes use their genomes during passage in mammals and ticks. Live imaging can further reveal in real time how arthropod-associated pathogens and symbionts
interact with their hosts, and point to ways to disrupt these interactions through genetic manipulation and mutagenesis. Molecular analysis is revealing the remarkable diversity of, and possible genetic exchange taking place within, this microbial community, which illuminates its capacity to adapt rapidly to new environments.
One hypothesis is that differential microbial gene expression may play a role in the pathogen’s adaptability to the markedly different environments in the mammalian host and tick vector. Gene expression was studied in cell lines that focused on the life cycle of A. phagocytophilum and represented the tick, the human endothelium, and the human granulocyte. Gene expression varied depending on the host cell line. For example, the outer membrane protein 1A is expressed quite well in mammalian cell lines, but not in the tick cell line. The outer membrane efflux protein, and the major surface protein 4, are seemingly expressed in tick cells, but not at all in mammalian cells. The heat shock protein is primarily expressed in mammalian cells, but less so in tick cells, perhaps reflecting the lower incubation temperature in the tick. However, a “housekeeping” gene required by the organism seems to be equally expressed in all cell lines, independent of their origin.
Mutational analysis is another technique to probe the biology of the pathogens and symbionts. Approximately 40 percent of the genome of A. phagocytophilum has no known function, and gene knockouts can reveal the function of those genes. Further explorations can be accomplished by overexpressing genes to obtain sufficient samples for biochemical and immunological characterization, and by studying promoters and gene regulation in vivo. Genetic analysis reveals that human pathogens are closely related to the symbionts of ticks, yet the symbionts do not infect humans or animals.
We have found that a single-plasmid construct encoding the Himar 1 transposase and a transposon could be used to mutagenize A. phagocytophilum when introduced into the bacteria by electroporation. The transposon is inserted randomly into the genome of the recipient bacteria by a “cut-and-paste” mechanism. This approach produces mutants that are then screened for their ability to replicate in tick cells, an endothelial cell line, or HL-60 cells (a human promelocytic cell line that can be differentiated into granulocytes). In experiments using two separate mutants of A. phagocytophilum with identified genomic insertion sites, we characterized the effect of gene disruption on the phenotype of the pathogen. One mutant that has an insert in the o-methyltransferase gene does not infect tick cells or even bind to them, but it does grow well in HL-60 cells. A second mutant has an insert into a large gene expressed only in the mammalian cells. This organism is able to grow in tick cells but no longer infects HL-60 cells, although it can infect laboratory hamsters.
Spotted-fever group rickettsiae cause reemerging TBDs. The incidence
of Rocky Mountain spotted fever caused by Rickettsia rickettsii has significantly increased since 2000 and has geographically shifted from the Rocky Mountain states to the South Central and southeastern United States. Pathogens in the spotted fever group occur in a wide range of ticks, including Dermacentor andersoni and D. variabilis (the Rocky Mountain wood tick and American dog tick, respectively), Rhipicephalus sanguineus (the brown dog tick), and Amblyomma maculatum (the Gulf Coast tick), whereas Ixodes scapularis carries a rickettsial symbiont that does not cause infection in animals and humans. We have successfully created a family of Himar 1 constructs that carry selectable and fluorescent markers to probe this group of pathogens. This has demonstrated that rickettsia, which were thought to only penetrate a tick’s midgut, and then somehow move into other organs through the hemolymph, actually travel from the midgut through tracheal air tubes throughout the tick’s body (Baldridge et al., 2007).
Recently, it was found that Rickettsia felis, which is transmitted by fleas, carries plasmids (Ogata et al., 2005). Since this initial finding, plasmids have been found in most rickettsia, including R. monacensis, R. peacockii, R. massiliae, R. amblyommii, R. hoogstraalii, and R. helvetica. Plasmids have thus far not been found in highly pathogenic species such as R. rickettsii and Rickettsia prowazekii, and the reasons for this true absence or lack of detection are unclear. Because plasmid-encoded genes are conserved across many species, the working hypothesis is that plasmids play an important role in the biology of rickettsia. Many genes on rickettsial plasmids are related to genes in other rickettsial species. However, there are genes on rickettsial plasmids that do not occur in other rickettsia but are found in unrelated bacteria. These findings suggest that rickettsial plasmids participate in horizontal gene transfer in these species. The genome of R. peacockii, which is the closest relative to virulent R. rickettsia, has a plasmid that encodes a cluster of genes related to the glycosylation island of Pseudomonas aeruginosa that are likely involved in phospholipid biosynthesis. The chromosome of R. peacockii encodes a gene important in phospholipid biosynthesis that has been mutated, and it is possible that the plasmid-encoded genes compensate for that mutation. Plasmids of different rickettsial species carry other genes that are only distantly related. For example, the parA proteins of R. peacockii and R. felis are most closely related to those of E. coli and Pseudomonas, whereas the small heat shock proteins Hsp-1 and -2 are tightly aligned with the rickettsial phylogenetic lineage.
The discovery of diverse plasmids in different Rickettsia species suggested their potential use as transformation vectors, a method that could greatly facilitate genetic analysis and accelerate understanding of pathogeneis. We developed a series of shuttle vectors starting with a recombinant
version of pRAM18, one of the plasmids from R. amblyommii, bearing fluorescent and selectable markers. The regions of the plasmid encoding the parA and DnaA genes, which are important in plasmid maintenance and replication, along with a selectable and fluorescent traceable marker, were subcloned into pGEM. Subsequently, screening of a pRAM18 library yielded evidence of another R. amblyommii plasmid, pRAM32, and the section containing the parA and DnaA genes was subcloned into pUC, another commercially available plasmid. This produced a family of constructs: the original recombinant, large-size pRAM18, its smaller derivatives, and the smaller pRAM32 construct. With all four of these constructs, transformants were obtained by electroporation of R. monacensis, R. montanensis, and R. bellii. The recombinant rickettsial pRAM18 plasmids were maintained as plasmids in rickettsial populations, as were the much smaller pGEM- and pUC-based constructs.
In conclusion, Munderloh noted that the Himar 1 transposase system is useful for random mutagenesis and gene knockouts in obligate intracellular bacteria, but it has proven not to be very efficient. Despite this shortcoming, Himar 1 transposon mutagenesis has enabled the study of gene function in organisms that traditionally have been difficult to manipulate genetically. In combination with plasmids used in complementation assays, existing constructs will enable investigators to make headway toward a functional genomic analysis of these bacteria. In this way, drugs targeting specific genes could be developed, or vaccine strains with attenuated virulence might be created that would generate a protective immune response without causing illness. The tools generated for rickettsial organisms could be useful for other pathogenic bacteria.
NATURAL HISTORY OF TICKS: EVOLUTION, ADAPTATION, AND BIOLOGY
Tom G. Schwan, Ph.D., M.S., Laboratory of Zoonotic Pathogens, National Institute of Allergy and Infectious Diseases
Ticks belong to the Phylum Arthropoda and the Class Arachnida. They are not insects. Although they have an exoskeleton and jointed appendages, they have eight legs, do not fly, and do not have a head, a thorax, or an abdomen. Ticks also differ vastly from most insects in that they are important vectors for many pathogens. Among those insects that feed on blood, such as mosquitoes, black flies, sand flies, tsetse flies, and fleas, only the adults—and often only females—feed on blood. That means that only adults can acquire an infectious blood meal from infected animals that serve as reservoirs for pathogens, such as rodents. In contrast, ticks are obligated
blood feeders at all stages of their life-cycle, which makes them adept at transmitting pathogens at various stages.
There are three families of ticks: Ixodidae, Argasidae, and Nutalliellidae. Ixodidae has 12 genera, while Argasidae has 4. Nutalliellidae consists of a single species. The ixodids are hard ticks and evolve through three stages: larva, nymphal, and adult. Argasids are soft ticks that also undergo multiple stages: larval, multiple nymphal stages, and mature adult. In contrast to hard ticks in which the adults feed only once, soft tick adults can feed multiple times. Not much is known about the evolution of ticks, as fossil ticks found in amber dating back 94 million years look like ticks of today (Klompen and Grimaldi, 2001).
Ixodid ticks have a life cycle of 1 to 3 years and are typically less able to fast and survive without a blood meal. In contrast, Argasidae have an expanded life cycle that may take many years to complete. These ticks are able to fast for long periods of time between blood meals. Ornithodoros parkeri females, O. tholozani nymphs, and O. moubata nymphs can live 10 to 11 years between meals (Schwan, unpublished) and may even outlive their vertebrate hosts by many years.
Ticks can be three-, two-, or one-host arthropods. For three-host ticks, larvae feed on a host, fall off, and molt into a nymph. The nymph then attaches to another host, feeds, and falls off, and finally the adult attaches to a third host and feeds. This group includes the hard tick I. scapularis, the vector for Lyme disease Borrelia. In the case of two-host ticks, some larvae and nymphs feed on a single host and then attach to a second host to reach adulthood. No North American ticks are two-host ticks. One-host ticks attach to a host as larvae, and then feed and mature to the adult stage on the same host. Rhipicephalus (Boophilus) microplus and relatives, which are cattle ticks, are classic one-host ticks. Deer, elk, and goats can have high numbers of Dermacentor albipictus, also a one-host tick, and hunters in North America often encounter these ticks.
There are approximately 870 species of ticks. Within this group, some ticks are widely distributed and feed on many different types of hosts, while other ticks are very host specific. I. scapularis, for example, is found throughout the eastern United States and feeds on 50 to 70 different hosts, while Argas monolakensis, an argasid, is found only on islands in Mono Lake, California, in high density and feeds only on the California gull. All ixodid ticks feed for long periods of time, while most argasids are fast feeders, although the larvae of some feed for many days. The latter is the case with A. monolakensis, which can exsanguinate gulls during the feeding.
In 1893, it was demonstrated that blood-feeding arthropods could be biological vectors of a pathogenic organism (Smith and Kilborne, 1893). Ticks are effective vectors and, sometimes, effective reservoirs. In some species, adult females can transmit the pathogens to their offspring as a
result of transovarial transmission. Infected ticks can also pass an organism from one development stage to subsequent stages in its life cycle, which is termed transstadial transmission. Because ticks feed on blood at every stage, live a long time, and can transmit pathogens to their offspring or next life cycle, they are capable of sustaining pathogens for long periods of time and they are exquisitely adapted to serve as reservoirs for pathogens and as effective transmitters. Moreover, all TBDs in North America are zoonoses—transmitted from animals to humans. However, the O. moubata tick can directly transmit B. duttonii, the pathogen that causes relapsing fever in East Africa, from person to person. In North America, O. hermsi ticks are nocturnal and fast feeding, and usually feed on people sleeping in tick-infested cabins.
Relapsing fever occurs in Africa, but it also occurs in the western United States, where it is underdiagnosed and underreported. The disease causes significant mortality in some regions of Africa, especially among pregnant women. Relapsing fever is usually not fatal in North America, but the risk of mortality increases for fetuses during pregnancy.
Knowledge Gaps and Research Opportunities
Schwan noted that the research needed in the future falls into two key areas:
-
The number and training of medical acarologists and tick biologists are declining, and scientists who do investigate TBDs often focus only on Lyme disease–related questions. Support for the training of tick biologists with wide-ranging interests and broad research portfolios are essential to ensure continued progress on the full spectrum of TBDs.
-
Field research on TBDs is particularly important: Analysis of ticks in the lab, using technologies such as polymerase chain reaction (PCR), is not enough.
WILDLIFE AND DOMESTIC HOSTS: THEIR IMPORTANT ROLES IN MAINTAINING AND AMPLIFYING PATHOGENS AND THEIR CHANGING DYNAMICS
Howard Ginsberg, Ph.D., U.S. Geological Survey and the University of Rhode Island
Ticks and their hosts can be influenced by environmental factors, and complex interactions influence the transmission of TBDs. This can be illustrated by Lyme disease in eastern North America, which is dependent
primarily on one vector and one pathogen, B. burgdorferi. Several factors influence the transmission of tick-borne illnesses in North America, including
-
Tick density, which affects the probability that both humans and reservoir hosts for B. burgdorferi—such as small rodents and birds—will be exposed to the bacteria.
-
Host factors, including the diversity of hosts and their competence as reservoirs for pathogens.
-
Spatial patterns, primarily the geographic distribution of I. scapularis ticks, the vectors for B. burgdorferi. That distribution can vary in the field meter by meter because of various microspatial factors.
-
Temporal patterns, primarily the length of the season during which ticks and their hosts are active.
Tick density influences the probability of exposures of both human and reservoir hosts of the pathogen. The probability that a host will be exposed to a pathogen that causes a tick-borne disease, given the density of ticks, can be expressed as Pe=1-(1-kv)n whereas Pe is the probability of being bitten by at least one infected vector, kv is the proportion of vectors infected with the pathogen, and n is the number of vector bites (Ginsberg, 1993). When the results are plotted for different rates of infection, it is an asymptotic curve, such that a 25 percent infection rate, which is common for nymphal ticks in endemic areas, means that by 5 to 10 tick bites, the probability of exposure is near one. On Fire Island, a barrier island that runs parallel to the south shore of Long Island, New York, most mice are bitten by 30 to 200 ticks per season. If a mouse is bitten by 100 ticks in an average season and a successful intervention decreases the number of bites to 20, there will still not be an effect on whether the mouse is going to be exposed to the pathogen. Therefore, trying to manage Lyme disease by developing strategies in the natural environment is difficult. Human risk, however, is lower because most individuals are bitten by only a few ticks in a given year. If the number of tick bites is lowered, then the probability of being exposed to the pathogen is lowered. Thus, managing the environment to minimize human–tick encounters is an easier approach than trying to manage the natural cycle.
A number of factors control the density of ticks. For example, the number of primary hosts in a region, such as deer, seems to determine the mean number around which the number of ticks fluctuates. However, broad fluctuations of the tick population occur from year to year. Tick populations in Westchester County north of New York City, Prudence Island in Narragansett Bay, and Fire Island off Long Island have similar yearly fluctuations, which suggests that whatever controls these fluctuations is occurring on
a regional scale (Ginsberg et al., 1998). However, weather alone apparently does not explain Lyme disease incidence as correlations with weather factors have variable results (McCabe and Bunnell, 2004; Ostfeld et al., 2006). The relationship between ticks and weather is complex and requires examination of conditions in the leaf litter where the ticks reside rather than simply measuring weather from an airport station. Furthermore, the amount of time that a tick spends below a certain humidity level affects both its survival and activity (Rodgers et al., 2007).
How the ticks interact with hosts can influence the number of ticks that acquire a pathogen from infected host animals, including the proportion of hosts that are infected, the reservoir competence of various species of hosts, and the distribution of larval and nymphal ticks on those hosts. Hosts, especially mammals and birds, vary significantly in their reservoir competence. Some domestic animals, such as dogs, are competent reservoirs for Lyme disease. However, domestic animals probably do not play a large role in the transmission cycle of Lyme disease because they spend much less time in the woods than do wild animals. There are exceptions: for example, on Monhegan Island, Maine, Norway rats are an invasive species that are competent reservoirs and run wild (Smith et al., 1993). Furthermore, the reservoir competence of hosts can vary significantly between laboratory and field studies (Table 4-1). In the laboratory, investigators study reservoir competence by putting infected ticks on animal hosts, or by injecting the animals with B. burgdorferi. A week or two later, the investigators put uninfected tick larvae on the now-infected animals and determine what percentage of the larvae acquire the bacteria. In field studies, by contrast, investigators put uninfected larvae on wild animals and determine how many acquire the infection. The results can vary between the laboratory and the field for reasons that are not well understood. For example, when robins were infected in the lab, they proved to be highly competent reservoirs for B. burgdorferi, with 82–92 percent of tick larvae acquiring the pathogen. By contrast, when uninfected tick larvae were placed on robins from the field, only 16 percent of the larvae acquired the bacteria (Richter et al., 2000; Ginsberg et al., 2005).
The reservoir competence of B. burgdorferi can also vary by geographic area depending on the diversity of the available hosts. Some of these host species are competent reservoirs and others are not. On Fire Island, uninfected I. scapularis larvae feed on a variety of host species—some good reservoirs and some not—with small rodents being the primary competent reservoirs. However, the community of hosts is not very diverse, so a high percentage of tick nymphs become infected. In a more ecologically diverse community, the variety of hosts for B. burgdorferi dilutes the impact of rodents as hosts (LoGiudice et al., 2003). That, in turn, affects the probability that people will acquire Lyme disease, although the effect is complex.
TABLE 4-1 Reservoir Competence of Selected Vertebrate Species for Borrelia burgdorferi (Using Xenodiagnostic Ixodes Scapularis Larvae)
|
Common Name |
Lab/Field |
Reservoir Competence |
|
|
(%) |
Source |
||
|
White-footed mouse |
L |
~75 |
Donahue et al. (1987) |
|
|
F |
89 |
Mather et al. (1989a) |
|
|
F |
56 |
Ginsberg et al. (unpub.) |
|
Meadow vole |
L |
~70 |
Markowski et al. (1998) |
|
|
F |
62 |
Markowski et al. (1998) |
|
Chipmunk |
F |
20 |
Mather et al. (1989b) |
|
White-tailed deer |
F |
1 |
Telford et al. (1988) |
|
American robin |
L |
92 |
Richter et al. (2000) |
|
|
L |
82 |
Ginsberg et al. (2005) |
|
|
F |
16 |
Ginsberg et al. (2005) |
|
Northern cardinal |
F |
9 |
Ginsberg et al. (2005) |
|
Gray catbird |
F |
0 |
Mather et al. (1989a) |
|
|
F |
4 |
Ginsberg et al. (2005) |
|
Eastern fence lizard |
L |
~7 |
Tsao et al. (2008) |
|
Five-lined skink |
L |
>20 |
Levin et al. (1996) |
|
Dog (beagle) |
L |
78 |
Mather and Ginesberg (1994) |
|
Norway rat |
F |
72 |
Smith et al. (1993) |
|
SOURCE: Ginsberg, unpublished. |
|||
For example, dilution works in the Northeast because small rodents, the primary host for B. burgdorferi, are very good reservoirs. In the South, lizards, the primary hosts for Ixodes scapularis, are not good reservoirs, so expanding the diversity of hosts in that region could actually increase the incidence of Lyme disease.
Vector diversity can also affect the likelihood of human disease. On Fire Island in the early 1980s, the American dog tick and the black-legged tick were the most common kinds of ticks. In the later 1980s and 1990s, the lone star tick, A. americanum, became more abundant, and the pathogens E. chaffeensis, E. ewingii, and R. amblyommii began to present risk for human disease (Mixson et al., 2006).
Spatial patterns include the geographic distribution of competent hosts and vectors. For I. scapularis, its distribution is common in the northern and much of the southern United States, but the cases of Lyme disease that are reported to the Centers for Disease Control and Prevention (CDC) are predominantly from the northern states. Thus, tick density alone does not determine these rates. One factor that might be important is the host competency. As noted above, mammals residing in the North are highly competent reservoirs for B. burgdorferi, while in the South the abundant lizards are not good reservoirs. A second spatial factor can be the genetic distribution of ticks. For example, one genetic lineage of I. scapularis ticks
occurs in both the northern and the southern United States, but numerous other lineages occur in the South but not the North (Beati, unpublished). Whether genetic lineages differ in host preference and vector competence is unknown.
Temporal patterns are also likely to be affected by geographic trends. In the Northeast, for example, adult I. scapularis lay eggs, and uninfected larvae hatch in midsummer and feed on infected hosts. Infected nymphs then emerge the following spring and take a blood meal, emerging as adults in the fall. That means the nymphs and larvae present in any given year represent different populations. Nymphs infect hosts with B. burgdorferi bacteria, which the larvae acquire feeding on these same hosts slightly later in the season; the transmission cycle is very efficient. If the seasonal cycles of larvae and nymphs overlapped even more, transmission could be less efficient. If nymphs emerged significantly earlier in the season and larvae considerably later, animal hosts could lose their infectivity during the interim period. Understanding the temporal relationship between these stages is therefore critical to understanding why Lyme disease is common in some areas and not in others. For example, understanding how a longer growing season affects the active season of various stages of the tick could inform how changing climate might affect human disease.
In a multilaboratory investigation of geographical patterns, standardized samples of host communities and the distribution of ticks on the hosts in four regions around the country are being collected to determine which genetic groups are present, and whether they correlate with ecological factors. Hypothesis testing and ecological modeling may help to determine why Lyme disease results in some areas with many infected ticks, while others have few. The results may shed light on how the distribution of Lyme disease might change in the future.
As discussed previously in the report, there are changing dynamics that will affect human disease. For example, tick distribution is expanding up the Hudson Valley in New York, into northern New Jersey, down into the Southeast, and into Illinois, and modeling suggests that the tick will expand further into Canada. Expanding tick distribution alone does not necessarily mean more human disease, but it is an important contributor. Another factor may be expanding host distribution. For example, if lizards expand north, the incidence of Lyme disease may decrease. Similarly, introduced species may influence disease transmission if they are competent hosts. Finally, changing active seasons of the ticks as a result of longer growing seasons may affect the transmission dynamics.
Knowledge Gaps and Research Opportunities
The key areas for future work Ginsberg noted include the following:
-
The effect of physical factors on the distribution and abundance of ticks.
-
The effect of changing climate on the distribution of vertebrate hosts of tick-borne pathogens.
-
The factors that influence the infectivity of those hosts in the field versus in the laboratory.
-
The influence of the length of the active season on tick phenologies, or life cycle events.
-
Geographic patterns in the genetic structure of tick populations.
-
Efficient targeting and integration of techniques for managing human exposure to ticks. For example, land-use planners and landscape architects could design communities and developments to reduce human exposure to ticks, even if they are relatively abundant. Such an approach would require a relatively modest investment.
DISCUSSION
King questioned the panel about what role pheromones may play in tick-borne disease. Munderloh noted that different species of ticks produce different types of pheromones for aggregation and for stimulation of mating. In addition, the composition of saliva is vastly different between I. scapularis and D. variabilis. Aggregation hormones play a major role in some ticks, such as D. variabilis, but play a lesser role in I. scapularis. It would be difficult because of the divergence of ticks to make generalizations. Ginsberg noted that there is active research on the use of pheromones to enhance tick management methods by attracting ticks to the pesticides, but nothing is commercially available.
Schutze questioned if freezing weather for longer than 3 days could affect the number ticks and thus the number of tick-borne illnesses that season. Ginsberg noted that laboratory studies suggest that the temperature has to be significantly below freezing before it has an effect. However, this may not be true in the field as ticks have adapted to finding places in the soil and under the surface that provide a measure of protection. Munderloh further noted that deep snow cover is protective because just underneath the snow cover some areas are barely below freezing.
Emerging infectious diseases seem to have a trajectory over time, where the incident rates increase and then plateau. Drawing from this observation, one participant questioned through pathogen, tick, or host dynamics what the limit will be on Lyme disease in the Northeast. Ginsberg noted that making any predictions will be difficult because Lyme disease is still spreading into new areas, and the increased reporting reflects this fact. The participant further questioned whether there is any long-time series of infection rates in hosts and ticks at multiple locations. Ginsberg noted that
infection rates vary significantly. On Fire Island, the infection rate can fluctuate from 4.5 percent to more than 30 percent from year to year. Although these are preliminary observations, he noted that the fluctuations may be related to reservoir host populations.
Walker asked if there is any effect of the prevalence of the pathogen in the ticks relative to the virulence of the pathogen. For example, this may be the case with D. variabilis that carry R. rickettsii and A. cajennense in South America that carry R. rickettsii in areas where the diseases are prevalent, but the tick numbers are relatively low. Schwan noted this question has not been well studied. Relapsing fever spirochetes may be detrimental to the tick if the ticks ingest a significant number of spirochetes, but this is anecdotal information. Higher environmental temperature increased the mortality of D. andersoni ticks when infected with R. rickettsii (Niebylski and Peacock, 1999).
Another participant questioned whether a host adapts over time to become an incompetent reservoir as its immune response adapts. For example, in a newly endemic area, the same species might be a good reservoir because it has not developed that type of immune response. Ginsberg noted that this might occur in nature and there may also be differences during the life cycle of the hosts, such that juveniles have a different reservoir competence than do adults. The participant further questioned if one of the reasons for the lack of overlap of the distribution of reported cases of Lyme disease and the distribution of the black-legged tick is the result of underreporting. Ginsberg noted that this could be one explanation but that the use of flagging techniques to collect ticks have found results that are comparable to CDC results about human cases.
COMPARATIVE MEDICAL IMPORTANCE OF A ONE-HEALTH APPROACH TO EMERGING TICK-BORNE DISEASES
Edward B. Breitschwerdt, D.V.M., College of Veterinary Medicine, North Carolina State University
A “One Health” approach recognizes the need for veterinarians, human health professionals, and environmental scientists to work together given the dynamic interface among people, animals, and the environment. This approach is increasingly important for zoonotic diseases, such as TBDs, which rely on animals as reservoirs. Numerous exchanges of knowledge, such as using animals as sentinels for human diseases, reveal the importance of a One Health framework for understanding TBDs. Use of this approach is likely to achieve advances in health care for the 21st century “by accelerating biomedical research discoveries, enhancing public health efficacy, expeditiously expanding the scientific knowledge base, and improving
medical education and clinical care” (http://www.onehealthinitiative.com/mission.php).
Animals, especially dogs, because of their close proximity to humans and the fact that they often present with similar signs of tick-borne disease, are often sentinels for human TBDs (Elchos and Goddard, 2003). For example, the symptoms of dogs infected either naturally or in the laboratory with R. rickettsii are nearly identical to those of humans with Rocky Mountain spotted fever. In the context of one medicine, veterinary medicine and human medicine can both provide key insights into TBDs. In a 2003 case study, a farm family’s dog died, despite treatment with antibiotics, after a delayed diagnosis of ehrlichiosis. Two weeks later the farm’s 46-year-old owner developed fever, headache, vomiting, and back pain. Because recognizing tick-borne illness in people is difficult, her family physician diagnosed muscle sprains and acute cystitis. The woman received two drugs, including a sulfonamides, which are contraindicated for the treatment of rickettsial diseases. She died 3 days later as a result of vasculitis. The CDC determined that she had had Rocky Mountain spotted fever. A week later the family’s second dog developed febrile illness and was quickly placed on doxycycline. A diagnosis of Rocky Mountain spotted fever was made based on seroconversion to R. rickettsia, as confirmed by the CDC (Elchos and Goddard, 2003).
In a second case study, an outbreak of Rocky Mountain spotted fever in the Bronx, New York City, in 1987 occurred most likely after a female tick was carried in by a dog that had traveled to an endemic area (Salgo et al., 1988). The tick, carrying R. rickettsii, presumably transmitted the pathogen transovarially to the next generation of baby ticks, which led to the outbreak of human disease. Today, as a result of improved acaricide products that are safer and more effective and tick-borne disease educational efforts by veterinarians, an urban outbreak is much less likely.
Surveillance and diagnostic advances in veterinary medicine continue to inform human tick-borne disease management and vice versa. Veterinarians routinely use a rapid, on-site surveillance test to determine whether dogs have been exposed to Ehrlichia sp., Borrelia burgdorferi (Lyme disease), or A. phagocytophilum; results from that testing have helped shed light on where human diseases caused by these pathogens can occur. Surveillance for E. canis, E. chaffeensis, and E. ewingii antibodies in dogs helped uncover that there is a high prevalence of brown dog ticks infected with and transmitting transmit E. canis in Arizona (Figure 4-1). The surveillance found a cluster of Ehrlichia in dogs in Minnesota and Wisconsin, which made sense after the CDC recently found an Ehrlichia muris–like infection in immunocompromised people in that region. Moreover, highly sensitive molecular tests for tick-borne organisms have helped to define the importance of coinfection and have clarified the pathogenesis and pathophysiology of TBDs.
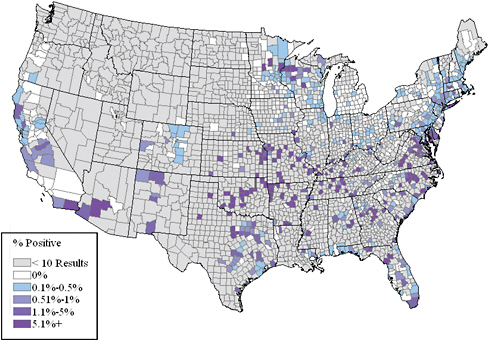
FIGURE 4-1 National prevalence data of Ehrlichia antibodies in dogs.
SOURCE: Reprinted from Veterinary Parasitology, 139, Bowman et al., Successful or currently ongoing parasite eradication programs, 138-148, Copyright 2009, with permission from Elsevier.
PCR analysis of peripheral blood samples has been useful in the diagnosis of Ehrlichiain dogs and humans. A recent report of E. chaffeensis in lemurs was confirmed by PCR (Williams et al., 2002).
Similar insights from naturally occurring infection in animals have enhanced understanding of human diseases due to other tick-borne pathogens. For example, A. phagocytophilum can cause illness in cats, dogs, horses, and humans in the same geographic area, with thrombocytopenia (low platelet count) the most consistent abnormality revealed by laboratory tests across species. Coinfection with B. burgdorferi is common because the same tick transmits both organisms. There is some experimental evidence of chronic infection of A. phagocytophilum in dogs (Scorpio et al., 2010), and this finding suggests the need to look at human illness caused by this pathogen. Notably, the discovery of human anaplasmosis occurred decades after veterinarians at the University of California identified the causative agent, then described as Ehrlichia. equi in the mid-1960s. Experimentally,
E. equi was transmitted to cats, dogs, and nonhuman primates to test how pathogenic the organism would prove to be across species lines. From these results a species of Ehrlichia was subsequently linked to human granulocytic ehrlichiosis (Bakken et al., 1994). This human pathogen was sequenced and found to be closely related phylogenetically to the E. equi reported decades earlier (Chen et al., 1994). Subsequent phylogenic studies resulted in reclassification of E. equi to the genus Anaplasma taxonomic unification of this agent with that of human disease, and speciation of A. phagocytophilum (Dumler et al., 2001).
Underscoring the need for a One Health approach, shortly after the discovery of human granulocytic anaplasmosis, researchers found by using DNA sequencing that dogs and horses with neutrophilic morula in Sweden were infected with a genetically identical segment of approximately 1,400 base pairs of the 16S rRNA gene to the Ehrlichia pathogen reported by Chen (Johansson et al., 1995). These results underscore the need for a One Health approach. As a result of sharing information across disciplines, across species, and across continents, granulocytic anaplasmosis was subsequently confirmed in cats and dogs in the northeastern United States and human anaplasmosis was reported in Sweden, Germany, Austria, and many other European and Asian countries.
Another opportunity for which veterinary research can provide insight into human disease is by using dogs as a naturally occurring model of disease. For example, in a recent study, whole blood and serum were collected from 731 dogs from a single veterinary hospital in Baxter, Minnesota. Clinical disease and a positive PCR test for A. phagocytophilum were more likely to occur in dogs previously infected with B. burgdorferi. The A. phagocytophilum and B. burgdorferi seroprevalence in dogs in this region was very high (55 percent, or 405 of 731 dogs tested) (Beall et al., 2008). For both of these pathogens, a dog was as likely to be healthy after having been exposed to and infected with these organisms than to present for disease manifestations. However, if these dogs had antibodies to both A. phagocytophilum and B. burgdorferi, then statistically the animals were clinically ill. Moreover, diagnostic documentation of A. phagocytophilum DNA in the blood was associated with illness. There is a high correlation between canine exposure to ticks that transmit A. phagocytophilum and B. burgdorferi and the likelihood that a person will be infected by ticks in the same environment. As veterinarians routinely screen for antibodies to these organisms, they can educate the client about the risk of TBDs in their pets and family members.
Bartonella is an alpha-proteobacterium and is phylogenetically related to other tick-borne pathogens in the genera Anaplasma, Ehrlichia, and Rickettsia. This may be the most important genus of bacteria infecting
both people and animals that scientists can study in the next decade. These organisms can be intraerythrocytic in humans, rodents, and cats alike. Unfortunately, unlike tick-borne organisms that have a tropism for only neutrophils (A. phagocytophilum or Ehrlichia ewingii) or macrophages (E. canis or E. chaffeensis, or primarily Bartonella), bacteria seem to invade erythrocytes, endothelial cells, and microglial cells as well as CD34 progenitor cells in the bone marrow. Currently, only circumstantial evidence supports tick transmission of Bartonella. Similarly, Bartonella underscores the need for One Health research because lice, fleas, sand flies, and possibly ticks can transmit pathogens in this genus of bacteria (Angelakis et al., 2010; Dietrich et al., 2010). In addition, people can become infected through animal bites, scratches, needle inoculation, and potentially through in utero transmission (Breitschwerdt et al., 2010; Oliveira et al., 2010). Gray squirrels, ground squirrels, and groundhogs can also serve as reservoirs for Bartonella—most likely different species of the bacterium. Meanwhile the spectrum of chronic human disease linked to this genus is expanding (Breitschwerdt et al., 2008, 2010a; Sykes et al., 2010).
Dogs and humans with bartonellosis share similar clinical manifestations, including culture-negative endocarditis (endocarditis without etiology), peliosis hepatis, bacillary angiomatosis, myocarditis, arthritis, encephalitis, immune-mediated thrombocytopenia, and immune-mediated hemolytic anemia (Chomel et al., 2009; Breitschwerdt et al., 2010a). Peliosis hepatis and bacillary angiomatosis occur in HIV-infected individuals and in dogs that are immunosuppressed due to cancer chemotherapy or immunosuppressive drug therapy (Yager et al., 2010). Furthermore, 32 percent of people with extensive contact with arthropods and animals are infected with sheep Bartonella, squirrel Bartonella, dog Bartonella, or cat Bartonella. Individuals who are not engaged in such contact show no reported infection using PCR analysis (Maggii et al., 2010).
Together Breitschwerdt noted that these findings for TBDs emphasize that collaboration among practitioners of veterinary and human medicine, along with overlapping surveillance systems, would be highly beneficial to both people and animals. Furthermore, there is a critical need to understand the role of vector-borne organisms as a cause of chronic disease in animals and humans. Finally, public education is important in preventing illness and death from acute infectious TBDs, such as anaplasmosis, ehrlichiosis, and Rocky Mountain spotted fever.
VARIATION OF BORRELIA SUBSPECIES: IMPLICATIONS FOR HUMAN DISEASE
James H. Oliver, Jr., Ph.D., Institute of Arthropodology and Parasitology, Georgia Southern University
Today, B. burgdorferi sensu lato complex includes 18 named and 1 not named yet spirochete species. Several of these subspecies are known human pathogens. In Europe, B. afzelii and B. garinii are among the most important genospecies causing human illness. In the United States, B. burgdorferi sensu stricto is considered to be the only causative agent of Lyme borreliosis until recently, when B. bissettii–like spirochetes were detected in people. Most of what is known about B. burgdorferi has been reported in the northeastern and midwestern states. For the southeastern United States, fewer studies have been conducted. To begin to investigate the differences in the southeastern United States, a number of working hypotheses of borreliosis have been proposed:
-
There is greater genetic diversity among B. burgdorferi s.l. in the southeastern United States.
-
Infectivity and pathogenicity of southern Borrelia vary more than northeastern strains.
-
Several “populations” of vector I. scapularis are distributed in the eastern and central United States. Tick behavior, life cycles, and Borrelia incidence vary among the populations and are determined by climate, local vegetation, suitable reservoirs or hosts, and the genetic profile of the Borrelia strains.
-
The largely non-human-biting I. minor and I. affinis are often infected with B. burgdorferi s.l. and serve as enzootic vectors of Borrelia.
-
Adult I. scapularis bite humans in the southeastern United States, but nymphs rarely do.
-
There is a wider diversity of vertebrate hosts of ticks in the southeastern United States, particularly reptiles, which probably serves to dilute incidence of B. burgdorferi in I. scapularis.
-
Birds are a major vehicle for long-distance transport of ticks and Borrelia.
-
Recent evidence suggests that one of the key determinants of spirochete–host association is the host complement system.
The 18 recognized genospecies of the B. burgdorferi sensu lato complex are listed in Table 4-2. Some have debated whether B. burgdorferi occurs in the southern states. Evidence in birds and animals of several subspecies
TABLE 4-2 Seventeen Recognized Genospecies of Borrelia burgdorferi Sensu Lato Complex
has found Borrelia in that area, including Georgia, Florida, South Carolina, and Missouri. These subspecies include B. americana, B. carolinensis, and B. bissettii. Furthermore, isolated genes from European Borrelia genotypes have been found in samples from birds and animals (Oliver, unpublished). In analyzing 112 subcultures from more than 300 isolates in those four states, 52 strains of B. burgdorferi have been found. Of those 52 strains, 15 had identity to the B31-type strain found in the northeast, while 37 had identity to other B. burgdorferi sensu stricto strains, including those from California and Europe (Oliver, unpublished). These data were based on two genomic loci, the 5S-23S intergenic spacer and the 16S rDNA.
An analysis of six genomic loci from B. burgdorferi strains in nymphal I. minor ticks found on a single Carolina wren in South Carolina underscores the great diversity of those strains. In nymphal ticks on a Carolina wren, two different genetic groups of B. americana were described. By pooling data from numerous I. minor larvae on the same bird, two other new subspecies of Borrelia were isolated (Oliver, unpublished). The fact
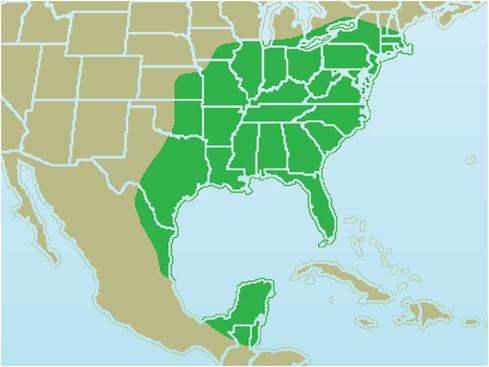
FIGURE 4-2 Birds are efficient transmitters of Borrelia strains over great geographic distances. For example, the range of Carolina wrens alone encompasses nearly half the United States.
SOURCE: Image courtesy of Ken Thomas, www.kenthomas.us.
that birds are efficient transmitters of Borrelia strains over great geographic distances promises to make these strains more widespread. The range of Carolina wrens alone encompasses nearly half the United States (Figure 4-2).
Among ticks that serve as vectors for B. burgdorferi, and among their animal hosts, there is significant genetic diversity. For example, we isolated 53 Borrelia strains in I. scapularis, 43 in I. dentatus, 27 strains in I. affinis, and 27 in I. minor. In addition, Borrelia strains were cultured from three primary rodents that serve as reservoirs of Borrelia in the south: Peromyscus gossypinus, the cotton mouse, had 70 strains; Sigmodon hispidus, the cotton rat had 26 strains; and Neotoma floridana, the eastern wood rat, had 35 strains (Oliver, unpublished).
There is also significant genetic diversity among Borrelia subspecies found to infect these bird and animal hosts. For example, in examining Borrelia isolates from five animals in South Carolina and Georgia—including a cotton rat, two cotton mice, a wood rat, and a downy woodpecker—the
researchers found both American and European strains, including sensu stricto, B. carolinensis, and B. garinii. In examining Borrelia isolates in 16 birds from St. Catherine’s Island, a small island south of Savannah, some birds carried only one genospecies while others carried three (Oliver, unpublished).
All these findings contradict recent dogma that Borrelia does not occur in the southern United States. In fact, Borrelia strains are spreading throughout the region. The question remains whether these strains cause human disease.
In summary, Oliver noted that more than 300 B. burgdorferi sensu lato have been isolated from Georgia, Florida, South Carolina, and Missouri. The largest numbers of isolates are B. burgdorferi, followed by B. bissettii from Georgia, South Carolina, and Florida, and then B. andersonii from Missouri, with B. carolinensis and B. americana recently described from Georgia and South Carolina. Most of the isolates have one genospecies, but 25 cultures contain more than one genospecies. Isolates from three sites in Georgia and two in South Carolina have genes from at least two European genospecies, B. garinii and B. afzelii. I. scapularis, I. affinis, and I. minor are the most common vectors of B. burgdorferi sensu lato in Georgia, Florida, and South Carolina.
Based on these findings and other research, Oliver noted a number of hypotheses for future research:
-
The infectivity and pathogenicity of Borrelia strains vary more in the southern states than in the northeastern states.
-
The behavior and life cycle of ticks and the incidence of Borrelia vary among tick populations, based on climate, seasonality, local vegetation, suitable reservoir hosts, and the genetic profile of the Borrelia strains.
-
I. minor and I. affinis are often infected with B. burgdorferi. Although those ticks rarely bite humans, they serve as enzootic vectors of Borrelia. That is, they keep Borrelia populations high in the environment. I. dentatus, also a common vector of B. andersonii, bites humans infrequently.
-
Unlike in the Northeast, I. scapularis nymphs rarely bite humans in the southern United States. However, contrary to belief, adult I. scapularis do bite humans in the South.
-
The southeastern United States has a wider diversity of vertebrate hosts, particularly reptiles, than does the northeastern United States, which may dilute the risk of human exposure.
-
The diversity of Borrelia associated with rodents is much lower than that associated with birds.
-
Genetically diverse strains of Borrelia often occur within the same individual tick or vertebrate host. Other investigators have shown that Borrelia has the recombination system needed for horizontal genetic exchange.
DISCUSSION
One participant questioned the accuracy of the tick distribution maps because many counties do not have an entomologist. Oliver noted that the distribution maps are incomplete because the collection of specimens is voluntary and relies on volunteers to send in samples. Another participant asked what a person should do given that the information on tick distribution is incomplete. Oliver noted that tick identification needs to be done. He said that most ticks do not transmit Borrelia and even if it is a species that does, the infected rate is low. Another participant noted that the lone star ticks A. americanum, do not carry B. burgdorferi, but may carry B. lonestari, which can cause Southern Tick-Associated Rash Illness (STARI). Oliver noted that STARI is not a well-understood area in the tick-borne disease field. He has found spirochetes on darkfield illumination, but has not been successful in culturing these.
Another participant noted that Columbia, Maryland, has one of the highest incidences of Lyme disease in the state. He asked the panel what communities can do to intervene in the tick life cycle, such as installing deer-feeding stations. Ginsberg noted that in highly endemic areas there is generally a need to provide an integrated approach that is tailored to the specific ecological conditions in the area. Four-poster feeding stations have been studied over a broad range of environments and have average effectiveness in controlling nymphal ticks; however, they will not do a complete job of prevention. There is a need to integrate different management methods, including education and environmental controls.
CONCLUDING THOUGHTS ON EMERGING INFECTIONS, TICK BIOLOGY, AND HOST–VECTOR INTERACTIONS
Lonnie King, D.V.M., M.S., M.A., College of Veterinary Medicine, Ohio State University
Because many TBDs are zoonotic, animal and human health experts urgently need to collaborate and to develop an integrated surveillance system that includes domestic animals, wildlife, ticks, and people. Wider and more effective surveillance could allow animals to serve as sentinels and surrogates for human risk and exposure to TBDs. Indeed, without robust diagnostics
and surveillance systems, TBDs are likely to remain underreported, and the true incidence and burden of these infections underappreciated.
In fact, surveillance conducted in Florida, Georgia, Missouri, and South Carolina suggests that the current map of the distribution of B. burgdorferi may be incomplete. Studies of the distribution, host, and diversity of isolates of B. burgdorferi in the Southeast also suggests the possibility that human risk in this region may be underrecognized and that the epidemiology is certainly poorly understood. The impact of regional differences in tick populations, hosts, habitats, and pathogens on human disease—and of genetically distinct subpopulations of those pathogens—deserve further study.
Although scientists know that ticks are coinfected with multiple microbes and pathogens, further research is needed to understand the roles and activities of these microbes and their interrelationships. Possible genetic exchange within this microbial community also needs further investigation, given that it could lead to new diseases.
Finally, comprehensive cross-disciplinary approaches to studying TBDs, and to improving prevention and treatment, are essential.
























