
3
Current Methods of Rangeland Assessment

The current theory and practice of rangeland assessment have a long history that is closely related to the ways that rangelands were used and studied. The nineteenth century was a period of exploration and development of the rangelands of the western United States. The need for systematic methods of rangeland assessment first became apparent when Jared Smith was sent by the U.S. Botanical Survey in 1895 to study the causes of the deterioration of western rangelands that had been widely reported in the late 1880s. He reported that:
The shortage of cattle all through the west is due to the fact that ranges were stocked up to the limit that they would carry during the series of exceptionally favorable years preceding the years of drought. Then followed the bad years when the native perennial grasses did not get rain enough to more than keep them alive. The cattle on the breeding grounds of the West and Southwest died by the thousands of thirst and starvation (Smith, 1896:322-323).
Such early investigations, however, were not based on a unifying science that could systematize the data collected to assess rangelands or relate the effects of livestock grazing to the rangeland deterioration that was evident in the late nineteenth century. The need for more thorough assessments was evident.
DEVELOPMENT OF CURRENT THEORY AND PRACTICE OF RANGELAND ASSESSMENTS
Between 1890 and 1905, 11 state agricultural experiment stations published 879 range management-related bulletins dealing with the control of weeds, pests, poisonous plants, soil moisture, fertility, conservation, rangeland inventory and analysis, water use, fencing, and other topics (Beetle, 1954). The U.S. Forest Service (USFS), which was formed in 1905,
Timeline of Rangeland Classification and Inventorying in the United States
|
Rangeland Use and Management |
Year |
Inventory |
|
Spanish settlers introduce sheep and cattle into California rangelands. |
1500 |
|
|
British settlers expand livestock grazing to more rangelands in the western United States. |
1800 |
|
|
Major increase in livestock on rangelands in Texas, Kansas, Nebraska, Oklahoma, and the Great Plains. |
1860 |
|
|
Overgrazing of the Great Plains and the Great Basin is accompanied by drought and severe erosion. |
1880 |
|
|
|
1895 |
Jared Smith conducts a survey of depleted western rangelands for the U.S. Botanical Survey. He reports that overgrazing and drought are responsible for widespread rangeland degradation. |
|
|
|
U.S. Department of Agriculture begins to publish reports on forage condition and grazing problems in the western United States. |
|
U.S. forest reserves are established, bringing part of western rangelands under the jurisdiction of the U.S. Department of Agriculture. |
1897 |
Surveys of forest reserves begin. |
|
|
1899 |
H.C. Cowles describes plant succession on the sand dunes of Lake Michigan. |
|
|
1900 |
State agricultural experiment stations begin to issue reports on the condition of rangelands and range management practices. |
|
Rangeland Use and Management |
Year |
Inventory |
|
U.S. Department of Agriculture establishes regulation of grazing on forest reserves. |
1901 |
|
|
The U.S. Forest Service (USFS) is formed and the National Forest system is created. |
1905 |
National forests are surveyed. |
|
|
1905 |
F. E. Clements begins adapting succession to Great Plains grasslands. |
|
|
1910 |
A. W. Sampson begins the first ecologically based range work of Utah. |
|
|
1917 |
J. T. Jardine develops the first scientific survey method. |
|
|
1923 |
A. W. Sampson introduces the use of succession as a way to assess grazing capacity. |
|
Dust Bowl ravages western rangeland and cropland. |
1930 |
|
|
|
1933 |
A. R. Standing proposes quantifying rangeland assessments by measuring volume of vegetation rather than estimating plant cover. |
|
The Taylor Grazing Act withdraws all remaining public land that is not under the jurisdiction of the USFS or other federal agencies into grazing districts under the jurisdiction of the Bureau of Land Management (BLM). BLM is charged with halting overgrazing and soil deterioration. |
1934 |
|
|
The Soil Conservation Service (SCS) is created. Its mandate is to inventory soil and water resources and to assist farmers and ranchers with ending erosion. |
1935 |
|
|
Rangeland Use and Management |
Year |
Inventory |
|
|
1936 |
The U.S. Senate issues a report on the condition of rangelands and the causes of rangeland deterioration. |
|
|
|
The interagency range survey technique is standardized in the late 1930s and used by BLM, SCS, and USFS to inventory rangelands. |
|
|
1938 |
J. E. Weaver and F. E. Clements publish Plant Ecology, extending successional concepts explicitly to rangeland ecosystems. Plant Ecology becomes the standard text for the emerging field of range management. |
|
|
1949 |
E. J. Dyksterhuis solidifies the use of successional stages as measures of rangeland condition by proposing the measurement of range condition as a departure from climax vegetation for a specific range site. Dyksterhuis' range site and condition method becomes the basis for SCS rangeland assessments. |
|
|
1950 |
USFS develops the Parker three-step method of rangeland assessment. |
|
|
|
BLM continues to use the interagency survey method. |
|
The Wilderness Act is passed. |
1964 |
|
|
The National Forest Management Act mandates multiple use and sustained-yield policies for national forest management. |
|
|
|
|
1968 |
R. Daubenmire describes habitat types, which become the basis for USFS forest and rangeland classifications. |
|
Rangeland Use and Management |
Year |
Inventory |
|
National Environmental Protection Act requires all federal agencies to write environmental impact statements on major federal actions. |
1969 |
|
|
The Endangered Species Act requires federal agencies to protect listed wildlife species. |
1973 |
|
|
The National Resources Defense Council v. Morton requires environmental impact statements on all local grazing programs administered by BLM. |
1974 |
|
|
The Resources Planning Act requires USFS to inventory national forests every 10 years. |
|
|
|
Congress passes the Forest and Rangeland Renewable Resources Research Act, which provides the information needed for Resources Planning Act implementation. |
|
|
|
|
1975 |
The first Resources Planning Act Assessment of resources, including rangelands on national forests, is published. |
|
Congress passes the Federal Land Policy and Management Act, which requires BLM to prepare an inventory of the resources on federal lands under BLM's jurisdiction. |
1976 |
|
|
The Soil and Water Resources Conservation Act is passed, requiring SCS to inventory soil, water, wildlife habitat, and related resources on nonfederal lands. SCS establishes the National Resources Inventory to carry out its mandate. |
1977 |
|
|
Rangeland Use and Management |
Year |
Inventory |
|
The Public Rangeland Improvement Act calls for improvements in soil quality, wildlife habitat, watersheds, and vegetation on federal rangelands and requires inventories of federal rangelands. |
1978 |
|
|
|
1979 |
BLM develops the soil-vegetation inventory method; the method is tested but is not formally adopted by BLM. |
|
|
1980 |
The second Resources Planning Act Assessment is published. |
|
|
1981 |
The first Resource Conservation Act Appraisal of soil and water resources on nonfederal lands based on the 1977 National Resources Inventory is published by SCS. |
|
|
1983 |
BLM adopts SCS range site and range condition procedures for assessing rangelands. |
|
|
|
The Society for Range Management recommends that SCS, BLM, and USFS adopt common terminology to classify rangelands and make ecological status ratings. |
|
|
1985 |
The National Resources Defense Council and the National Audubon Society assemble data on the rangelands under BLM's jurisdiction and report that many rangelands are in unsatisfactory condition. |
|
|
1987 |
The second Resources Conservation Act Appraisal of soil and water resources on the 1982 National Resources Inventory, is published by SCS. |
|
Rangeland Use and Management |
Year |
Inventory |
|
|
1989 |
The Society for Range Management assembles data from SCS, BLM, and USFS in an attempt to make a national assessment of rangelands. The society reports that the data available for federal rangelands are not adequate for a national assessment. The USFS publishes an assessment of the range resources of the United States. |
|
|
1990 |
The third Resources Planning Act Assessment is published. |
recognized the need to develop a scientifically credible and economically feasible method of surveying rangelands to carry out its mandate.
Since the goal of most of the early rangeland professionals was to provide high-quality livestock forage, the techniques and systems they developed for rangeland assessments concentrated on the effects of livestock grazing on forage production. The first formal attempt to develop a scientific rangeland survey method was made by James L. Jardine on the Conconino National Forest in 1910 (Chapline and Campbell, 1944).
Early Development Of Survey Methods
Jardine's range reconnaissance method involved a careful visual examination of the rangeland to provide a written record of the rangeland's resources. He recorded the following data: (1) a topographic map showing watering places, roads, fences, and cabins; (2) a classification of the rangeland into 1 of 10 grazing or vegetation types; (3) the percentage of the rangeland covered by each forage species; (4) a descriptive report of each grazing or vegetation type, including the suitability of each type for each kind of grazing animal; (5) a map of the timber; and (6) samples of the major species present on the rangeland (Jardine and Anderson, 1919).
Jardine's survey method was highly credible in its time, but it had several shortcomings regarding forage availability estimates. For example, it was based on estimates of the ground cover of each species rather than on direct measurements of the volume or weight of the forage pro-
duced by each plant species, and it therefore did not give an accurate measurement of productivity or yield.
Standardization of Rangeland Surveys
In 1933, Standing introduced the concept of using measured volumes of vegetation rather than visual estimates of cover (Standing, 1933). During the 1930s, other modifications were made to the Jardine method, and these were finally standardized as the interagency range survey technique used by the Bureau of Land Management (BLM) and USFS. Although more quantitative than the original reconnaissance method, the interagency survey depended heavily on palatability factors and other subjective criteria for estimating forage production or carrying capacity. This method assessed, almost exclusively, forage production and livestock carrying capacity. Few if any data were collected on soil conditions, wind and water erosion, or other factors that would allow a more comprehensive evaluation of rangelands. More important, the method was not linked to any theoretical base that suggested how the forage composition data that were collected could be interpreted as indicators of ecological conditions on rangelands. Forage production, rather than the state of rangeland ecosystems, was evaluated.
New Theoretical Foundation for Rangeland Surveys
At the same time that Jardine was developing his method for evaluating rangelands, ecologists were developing theories of community dynamics (how plant communities develop and change) that would provide the foundation for new methods for evaluating rangelands.
SUCCESSION AND CLIMAX COMMUNITIES
F. E. Clements of the University of Nebraska, Lincoln, was extremely influential in the study of succession in the Great Plains grasslands. His numerous publications on plant succession and ecology formed a major source of information for resource managers. The textbook Plant Ecology, which Clements wrote with his colleague J. E. Weaver (Weaver and Clements, 1938), became a standard in the field. Students from the ''Nebraska school of ecology'' such as E W. Albertson, E. J. Dyksterhuis, A. W. Sampson, and L. A. Stoddart became leaders in the young science of rangeland management and brought the Clementsian model of community change into the new field. The Clementsian model dominated much of the early literature in the field.
Clements developed a theory of vegetation dynamics and a quantita-
tive method to test his theory. To Clements, the climax theory rested on the assumption that vegetation could be classified into formations that represented a group of plant species that acted together as if they were a single organism. He wrote, ''As an organism, the formation arises, grows, matures, and dies.... each climax formation is able to reproduce itself, repeating with essential fidelity the stages of its development" (Clements, 1916:3).
The climax formation was "the climax community of a natural area in which the essential climatic relations are similar or identical" (Clements, 1916:126). (A climax community is the assemblage of plant species that most nearly achieves a long-term steady state of productivity, structure, and composition on a given site [Tueller, 1973].) Clements believed that all successional units within a climatic region developed along one linear path toward a plant community climax that was determined by climate (a climatic climax community). Thus, within a climatic region, a group of plant species would be identified as the climax vegetation, and all sites within that region could be compared with the climax plant species to determine where in the successional path the site was. This theory of vegetation dynamics has been referred to as the monoclimax theory.
Clements' method of vegetation analysis involved the use of permanently located quadrats (a plot, usually rectangular, used for ecological and population studies). The species of vegetation in the quadrat was carefully plotted on a map. Changes in vegetation were determined by periodically replotting on a map the species that were present.
The concept of successional change in rangeland ecosystems was to become the fundamental basis of the methods used today to inventory and classify rangelands. Rangelands would be classified on the basis of differences in climax plant community composition and assessed on the basis of the divergence of the current plant composition from the climax plant community composition.
SUCCESSIONAL STAGES AND RANGELAND ASSESSMENT
Sampson (1917) provided what was perhaps the first published reference on the utility of successional stages in rangeland assessment. Then, in 1923, Sampson wrote about the need to move from the old method of determining grazing capacity, which used palatability factors and visual estimates of forage composition, to a new method based on observation of the succession of conspicuous vegetation, that is, the replacement of one set or type of plants by another (Sampson, 1923).
Sampson studied community development in the Watasch Mountains in Utah and classified four developmental stages: the climax herbaceous stage, the mixed grass and weed stage, the late weed stage, and the early
weed stage. Although Sampson acknowledged that the Watasch Mountain climax species were not found everywhere, he noted that the character of growth and the habitat requirements of the plants of the different stages were generally the same on native pasturelands. In describing forage production during these four stages, he noted that the climax and the mixed grass and weed stages produced the most forage in terms of quantity and quality (Sampson, 1923).
Sampson noted that the use of successional units to develop a rational grazing plan presumed a detailed knowledge of the successional stages in the development of the vegetation (Sampson, 1923). To obtain this information, he recommended the use of quadrats. However, the great amount of tedious work involved in the mapping and the subsequent synthesis of the data led Sampson to recommend that the person working in the field record the percent cover of all plants of each species within each of the 100 cells that divided the chart quadrat rather than mark the specific location of each plant within each cell. This cover estimate was then multiplied by the palatability of the cover to determine forage yield. Sampson's work was instrumental in bringing successional theory and practical grazing management together.
SUCCESSIONAL STAGES AS CONDITION CLASSES
Sampson's ideas spawned much research into using successional stages as indicators of the status of rangelands. A number of rangeland scientists experimented with methods that could be used to determine the relationship of successional stages to rangeland condition in, for example, Colorado (Hanson et al., 1931), Kansas (Albertson, 1937), Nebraska (Weaver and Fitzpatrick, 1932), North Dakota (Hanson and Whitman, 1938; Sarvis, 1920, 1941), and the intermountain region (Sampson, 1919, 1923).
In 1949, E. J. Dyksterhuis published a landmark paper that was to solidify the contribution of successional theory to the assessment of rangelands. Dyksterhuis refined the climatic climax community described by Clements (1916), proposing that different climaxes coexist as a function of soil or topographic or geographic differences within a similar climate. Dyksterhuis defined those areas that support a unique climax community as a range site. Each site—defined by its climax plant community, soil, and climatic environment—would support a characteristic assemblage of plants, and this vegetation would persist unless it was disturbed by grazing, fire, drought, or other factors. Vegetation would develop toward this climax plant community through successional processes once disturbances (wind, drought, fire) ceased. Grazing drove the plant composition toward the early stages of succession, whereas natural successional pro-
cesses drove plant composition toward a climax community. By adjusting the grazing pressure or the duration or season of use, rangeland managers could maintain rangelands at any stage of succession.
Dyksterhuis proposed a quantitative system for assessing whether a rangeland was at an early or late stage of succession by analyzing the behaviors of three classes of plant species: decreasers, increasers, and invaders. As livestock grazing drove the plant composition toward earlier stages of succession, certain plants were thought to decrease in abundance. These decreasers were replaced by other plants that initially increased in abundance. Those increaser plants were thought to decrease in number and abundance if grazing pushed the plant composition to even earlier stages of succession. The plants that replaced the increasers were called invaders. The successional stage that the rangeland was in could then be determined by what proportion of the vegetation, measured by percent composition by weight, was decreasers, increasers, or invaders. If most of the plants were decreasers, the rangeland was thought to be in a late successional stage; if most plants were invaders, the rangeland was considered to be in a very early stage of succession.
Dyksterhuis also proposed that the condition of rangelands improved as succession progressed. Later successional stages were thought to provide better forage and to be more stable and productive plant communities. The condition of a rangeland could therefore be determined by the climax plant community of the site. The greater the proportion of increasers or invaders, the poorer the condition. The greater the proportion of decreasers, the better the condition.
ADOPTION OF THE SUCCESSION-RETROGRESSION MODEL BY FEDERAL AGENCIES
Dyksterhuis's use of successional stages as the measure of the condition of rangelands had great appeal. His concept not only proposed a systematic way of evaluating the condition of rangelands but also explained the effects of grazing on rangeland vegetation and provided the basis for changes in grazing management. Estimations of livestock carrying capacity were linked to range sites, condition classes, and successional stages. By 1950, the measurement of range condition (Soil Conservation Service [SCS]) as the degree of departure from climax plant community (SCS) vegetation of a defined range site and the succession-retrogression model of rangeland development became the standard concept in U.S. rangeland management. All major inventory and classification methods in use today are modifications of that basic concept.
The concept was adopted to varying degrees by the USFS, BLM, and
SCS, the agencies with the most responsibility for rangeland management in the United States. Changes in terminology and interpretation since 1950 have resulted in divergences between the site classification definitions and the rangeland inventory methods used by the different agencies.
APPLICABILITY OF THE SUCCESSION-RETROGRESSION MODEL
Even as the succession-retrogression model was accepted by rangeland scientists and institutionalized in the federal management agencies (SCS, BLM, USFS), other community ecologists began to question the validity of the concept of climax community itself.
This debate was missing from the first editions of Stoddart and Smith's (1943) and Sampson's (1952) range management textbooks (Smith, 1989) and is still missing from the later textbooks of Stoddart and colleagues (1975) and Heady (1975) and the most recent range research methods book edited by Cook and Stubbendieck (1986). The books and reference papers leading to the development of range site (SCS) and range condition (SCS), however, make no mention of papers by Cain, Egler, or Gleason that questioned the successional model (Smith, 1989).
Within the range science literature, investigators criticized the subjective nature of habitat type (West, 1982) and range site (Laycock, 1989) classifications, yet the Range Inventory Standardization Committee of the Society for Range Management recommended governmentwide use of the ecological site classification system (Society for Range Management, Range Inventory Standardization Committee, 1983), which was based on the same community ecology theories of previous classifications. New developments in community ecology including the analysis of community structure and causal factors done by multivariate techniques (statistical analysis of the interaction of multiple causes of rangeland change) and new community ecology ideas, like the threshold concepts of Friedel (1991), have not been incorporated into the methods used to inventory, classify, or monitor rangelands.
LINKS BETWEEN OTHER BRANCHES OF ECOLOGY AND RANGELAND SCIENCE
New developments in ecological research have had an influence on specialization and research within the field of range science, but this influence on the diversity of range science research has not yet been transferred to a diversification of the measures used to inventory and monitor rangeland. The fundamental concepts underlying the rangeland classifi-
cation and inventory methods of all of the federal agencies are based on those proposed by Sampson, Clements, and Dyksterhuis in the first half of the twentieth century.
CURRENT AGENCY RANGELAND ASSESSMENT THEORY AND PRACTICE
All federal agencies measure range condition (SCS) or ecological status (USFS and BLM) as the degree to which the vegetation of a site is different from the climax plant community or potential natural community characteristic of that or similar sites. SCS, USFS, and BLM have adopted systems that use (1) ecological site (BLM) or range site (SCS) as the landscape subdivision on which the analysis is made, (2) climax plant community (SCS) or potential natural community (USFS and BLM) as the standard against which range condition (SCS) or ecological status (USFS and BLM) is judged, and (3) succession and retrogression models as the primary means of explaining the ways that rangelands change.
When and how these agencies use range site (SCS) or ecological site (USFS and BLM) and how a rangeland is given a range condition (SCS) or ecological status (USFS and BLM) rating vary. SCS assists nonfederal landowners with developing and implementing conservation plans to protect soil, water, and other natural resources on their rangelands and conducts the National Resources Inventory. The methods used to classify sites and the soils on those sites are standardized at the national level. The SCS National Range Handbook (U.S. Department of Agriculture, Soil Conservation Service, 1976) prescribes the procedures that should be used by agency employees. Each state SCS office standardizes the methods used within that state to evaluate rangelands.
USFS and BLM have responsibility for managing federal lands for multiple uses on a sustained-yield basis and for maintaining a data base of the ecological status (USFS and BLM) of the lands under their jurisdiction. The site classification and evaluation methods used by BLM are similar to those used by SCS and are standardized at the national level. The USFS site classification and evaluation methods have recently been standardized at the national level in newly released manuals (U.S. Department of Agriculture, Forest Service, 1991b).
The responsibility for developing and implementing the ecological type (USFS) classification system in USFS has been assigned to regional foresters and forest and range experiment station directors. These individuals also have responsibility for correlating ecological type (USFS) descriptions across regional boundaries and between and among other agencies (U.S. Department of Agriculture, U.S. Forest Service, 1991b).
Rangeland Reference Terms
Three federal agencies (the Bureau of Land Management [BLM], the U.S. Forest Service [USFS], and the Soil Conservation Service [SCS]) evaluate and classify rangelands, but the techniques and evaluation criteria vary somewhat. The following is a comparative analysis of the definitions each agency uses.
|
Terms of Reference |
Agency |
Definition |
|
Status Rating |
||
|
Ecological status |
BLM, USFS |
Four classes used to express the degree to which the makeup of the present vegetation reflects the potential natural community. The class or rating, percentage of vegetation present in a potential natural community, is as follows: potential natural community, 76 to 100 percent; late seral, 51 to 75 percent; midseral, 26 to 50 percent; early seral, 0 to 25 percent. |
|
Range condition |
SCS |
Four classes used to express the degree to which the makeup of the present vegetation reflects the climax plant community. The class or rating, percentage of vegetation present in a climax plant community, is as follows: excellent, 76 to 100 percent; good, 51 to 75 percent; fair, 26 to 50 percent; poor, 0 to 25 percent. |
|
Site Classification |
||
|
Range site |
SCS |
A distinctive kind of rangeland that differs from other kinds of rangelands in its ability to produce a characteristic natural climax plant community. |
|
Ecological type |
USFS |
A category of land with a unique combination of potential natural community, soil landscape features, and climate; it differs from other ecological types in its ability to produce vegetation and respond to management. |
|
Terms of Reference |
Agency |
Definition |
|
|
||
|
Ecological site |
BLM |
A kind of land with a specific potential natural community and specific physical site characteristics; it differs from other kinds of land in its ability to produce vegetation and respond to management. |
|
Trend |
SCS, USFS |
Trend is described as up, down, or not apparent (also static or stable). Up represents a change toward a climax of potential natural community; down means a change away from a climax or potential natural community; not apparent means there is no recognizable change. |
|
Apparent trend |
SCS, BLM |
A judgment of trend based on a one-time observation. It includes consideration of such factors as plant vigor; abundance of seedlings and young plants; accumulation or lack of plant residues on the soil surface; and soil surface characteristics including crusting, gravel pavement, pedestaled plants, and sheet or rill erosion. |
|
Benchmark Plant Communities |
||
|
Climax plant community |
SCS |
The natural plant community that would be found on a range site in the absence of abnormal disturbances and physical site deterioration. It includes only native plant species. |
|
Potential natural community |
USFS, BLM |
The biotic community that would become established if all successional sequences were completed without interferences by humans under the present environmental conditions. It may include naturalized nonnative species. |
Site Classification
SCS uses the term "range site" to classify different rangelands. BLM has adopted site classifications that are similar in concept to those of SCS. USFS uses the term "ecological type" to classify its rangelands. All three agencies classify rangelands into different types on the basis of the kinds and amounts of plants expected in the climax plant community (SCS) or potential natural community (USFS and BLM) thought to be characteristic of that type of rangeland.
RANGE SITE CLASSIFICATION
SCS pioneered the use of range sites for rangeland classification and has used the concept fairly consistently since the 1940s. The SCS National Range Handbook (U.S. Department of Agriculture, Soil Conservation Service, 1976) defines a range site (SCS) as a specific area that is "capable of supporting a native plant community typified by an association of species that differs from that of other rangeland sites in the kind or proportion of species or in total production" (Section 302.1). Since a range site (SCS) is defined as a native plant community, no introduced or exotic species can be considered part of the climax plant community used to define the range site (SCS). This restriction of the definition to native plants is the only major difference that distinguishes range site from the site classifications used by USFS and BLM.
SCS considers each site to be the product of all the environmental factors responsible for its development, including soils, vegetation, topography, climate, and fire. Soil surveys are particularly useful in site classification. Soil surveys classify soils with similar properties into mapping units. The characteristics that are used to classify soils are primarily those that can be measured in the field such as color, arrangement of horizons, pH, texture, and other morphological features. From knowledge of the mapping unit that contains the soil being classified, SCS soil scientists can estimate such soil attributes as the depth of the soil influenced by organic matter, the mineral or chemical content of soil horizons, the rate at which the soil takes up water, the water storage capacity of the soil, the soil's vulnerability to erosion, and the soil's fertility. The differences in soil properties described in soil surveys are important elements in classifying rangelands into range sites (SCS).
For each site, the climax plant community composition is defined as that which existed before human influence. In the absence of abnormal disturbances that upset ecological processes or physical deterioration of the site, the unique interaction on a given site is thought to support a plant community characterized by plant species that differ from those in
the plant community found on another site in terms of the kind or proportion of species or total annual vegetative production.
Range sites are mapped and correlated by matching soils and climate zones with what is determined to be the characteristic climax vegetation. Data for this purpose are derived from many sources, including the following:
-
evaluation of the vegetation and soils on rangelands that have been protected from disturbance for long periods of time;
-
comparison of areas that are used to various degrees by livestock with similar ungrazed areas;
-
evaluation and interpretation of research dealing with natural plant communities and soils;
-
review of early historical and botanical literature; and
-
prediction of climax vegetation on the basis of information gathered from areas with similar soils and climates.
SOILS AND CLIMATE Soils and climates that result in the same climax vegetation and annual biomass production are considered to make up the same range site (SCS). Plant composition, measured by the weight of biomass produced annually by each species, is the key descriptor of a range site (SCS). The range site (SCS) usually remains the same as long as the soft and climate remain unchanged. If the soil has been changed by erosion or some other factor so that the changed soft in combination with the site's climate cannot support the growth of the characteristic climax vegetation, then a new range site (SCS) must be defined.
Individual sites are identified and differentiated from others on the basis of specific criteria. The criteria are
(1) significant differences in the species or species groups that are ecological dominants in the plant community; (2) significant differences in the proportion of species or species groups that are ecological dominants of the plant community; and (3) significant differences in the total annual production of the plant community (U.S. Department of Agriculture, Soil Conservation Service, 1976:Section 302.6).
Table 3-1 provides a comparison of the vegetation found on three different soft types in the same county in Utah and illustrates how plant community data are used to arrive at rangeland site definitions (Shiflet, 1973). All three soils support essentially the same plant community in terms of species composition. All sites are dominated by bluebunch wheatgrass (Agropyron spicatum), with only minor differences in the other components. However, there was a statistically significant difference in total production between the Manila and the Broad soils, with the Manila soil being approximately 17 percent more productive than the Broad soil.
Table 3-1 Average Production and Composition of Vegetation Produced on Three Soil Types in Box Elder County, Utah
|
|
Middle Soila |
Broad Soilb |
Manila Soilc |
|||
|
Major Species |
Production (kg/ha) |
Composition (percent) |
Production (kg/ha) |
Composition (percent) |
Production (kg/ha) |
Composition (percent) |
|
Bluebunch wheatgrass (Agropyron spicatum) |
1,650* |
85 |
1,462+ |
82 |
1,833* |
88 |
|
Sandberg bluegrass (Poa secunda) |
32 |
2 |
58 |
3 |
24 |
1 |
|
Balsamroot (Balsamorhiza sagittata) |
50 |
3 |
34 |
2 |
7 |
T |
|
Cheatgrass (Bromus tectorum) |
9 |
T |
7 |
T |
1 |
T |
|
Yellowbrush (Chrysothamnus viscidiflorus var. lanceolatus) |
32 |
2 |
64 |
4 |
36 |
2 |
|
Big sagebrush (Artemisia tridentata) |
29 |
1 |
32 |
2 |
30 |
1 |
|
Bitterbrush (Purshia tridentata) |
44 |
2 |
— |
— |
1 |
T |
|
Other species |
92 |
5 |
116 |
7 |
140 |
8 |
|
Total |
1,938*,+ |
100 |
1,773+ |
100 |
2,072* |
100 |
|
NOTE: Data are based on 30 observations of 10 subplots each. Reading across columns, production values followed by the same symbol (* or +) were not significantly different at the 5 percent probability level. T, trace percentage (less than 0.5 percent); —, did not occur or does not apply. a Loamy-skeletal, mixed, mesic Calcic Haploxeroll. b Loamy-skeletal, mixed, frigid Calcic Argixeroll. c Fine, Montmorillonitic, frigid Typic Argixeroll. SOURCE: Adapted from T. N. Shiflet. 1973. Range sites and soils in the United States. Pp. 26-33 in Arid Shrublands: Proceedings of the Third Annual Workshop of the United States/Australia Rangeland Panel, D. H. Hyder, ed. Denver: Society for Range Management. |
||||||
Assuming that a 15 percent difference were large enough to affect grazing management, the plant community that grows on Manila soils would be classified as a separate site.
An example of how differences in soils, production, and plant composition interact to influence the determination of rangeland site can be seen in three rangelands that were studied for 10 years in a project described by Williams and Hugie (1966) (Table 3-2). The only major difference in the environments of the three study locations was the soil. Production from the Hoelzle and Bancroft soils did not differ significantly. On average, however, the plant communities on these soils were approximately 35 percent more productive than those that grew on Goodington soils. This difference was statistically significant and large enough to require different management practices (Shiflet, 1973). The plant community that grew on Goodington soft would therefore be separated from those that grew on the other two soft types because of its lower productivity. Plants that grew on the Hoelzle and Bancroft soils did not differ in production, but they did differ in species composition. Idaho fescue (Festuca idahoensis) was a significantly higher producer in the plant community that grew on Hoelzle soils than it was in the community that grew on Bancroft soils, even though it was the most important herbaceous species on both soils. Another difference in the two plant communities was within the shrub component. Big sagebrush (Artemisia tridentata) was the major shrub in the community growing on Hoelzle soil but did not occur at all in the plant community growing on Bancroft soils. On the other hand, three-tip sagebrush (Artemisia tripartita) accounted for only 2 percent of the production of the vegetation on Hoelzle soils but was the most important shrub on the Bancroft soils, contributing 17 percent of the total production. On the basis of the lower productivity of the plant community growing on Goodington soils and differences in species composition and proportion of species between the communities growing on the Hoelzle and Bancroft soils, Williams and Hugie concluded that all three were unique and represented three distinct range sites (SCS) (Shiflet, 1973).
TOPOGRAPHY Topography, too, can play an important role in distinguishing between range sites (SCS). Features such as the slope of the land, the direction the sloping land faces and whether the land is located at the top or bottom of the slope affect runoff and delivery of water, evaporation, temperature, and other factors that influence the kinds and amounts of plants that grow on a site. Often, soils that are quite similar in many respects support different vegetation because of the influence of topography and are classified as different range sites (SCS). Table 3-3 illustrates the effects of exposure on two study areas located on opposite sides of a hill in southern Idaho. The soils on the two sites were very similar. There were only minor differences in the structures of the two plant communi-
Table 3-2 Average Production and Composition of Vegetation Produced on Three Soils in Blaine County, Idaho
|
|
Goodington Soila |
Hoelzle Soilb |
Bancroft Soilc |
|||
|
Major Species |
Production (kg/ha) |
Composition (percent) |
Production (kg/ha) |
Composition (percent) |
Production (kg/ha) |
Composition (percent) |
|
Idaho fescue (Festuca idahoensis) |
187 |
24 |
430 |
39 |
257 |
24 |
|
Bluebunch wheatgrass (Agropyron spicatum) |
62 |
8 |
121 |
11 |
159 |
15 |
|
Sandberg bluegrass (Poa secunda) |
144 |
18 |
149 |
13 |
91 |
9 |
|
Bottlebrush squirreltail (Sitanion hystrix) |
94 |
12 |
41 |
4 |
— |
— |
|
Prairie June grass (Koelaria cristata) |
13 |
2 |
6 |
1 |
84 |
8 |
|
Narrowleaf pusseytoes (Antennaria stenophylla) |
43 |
5 |
— |
— |
— |
— |
|
Longleaf phlox (Phlox longifolia) |
31 |
4 |
19 |
2 |
26 |
|
|
Hawksbeard (Crepis acuminata) |
— |
— |
9 |
1 |
47 |
4 |
|
|
Goodington Soila |
Hoelzle Soilb |
Bancroft Soilc |
|||
|
Major Species |
Production (kg/ha) |
Composition (percent) |
Production (kg/ha) |
Composition (percent) |
Production (kg/ha) |
Composition (percent) |
|
Balsamroot (Balsamorhiza sagittata) |
— |
— |
8 |
1 |
— |
— |
|
Other annuals |
55 |
7 |
38 |
3 |
11 |
1 |
|
Big sagebrush (Artermisia tridentata) |
25 |
3 |
67 |
6 |
— |
— |
|
Three-tip sagebrush (Artemisia tripartata) |
11 |
1 |
21 |
2 |
175 |
17 |
|
Desert rabbitbrush (Chrysothamnus viscidiflorus) |
14 |
2 |
17 |
2 |
27 |
3 |
|
Other species |
114 |
14 |
177 |
15 |
166 |
16 |
|
Total |
793* |
100 |
1,103+ |
100 |
1,043+ |
100 |
|
NOTE: Data are based on 10 annual observations of 20 subplots each. Reading across columns, production values followed by the same symbol (* or +) were not significantly different at the 5 percent probability level. T, trace percentage (less than 0.5 percent); —, did not occur or does not apply. a Fine, Montmorillonitic, frigid Typic Palexeroll. b Has been eliminated as a separate soil series. c Fine-silty, mixed, frigid Calcic Argixeroll. SOURCE: Adapted from T. N. Shiflet. 1973. Range sites and soils in the United States. Pp. 26-33 in Arid Shrublands: Proceedings of the Third Annual Workshop of the United States/Australia Rangeland Panel, D. H. Hyder, ed. Denver: Society for Range Management. |
||||||
Table 3-3 Forage Production, Composition, and Frequency of Vegetation Produced on Two Exposures of a Silt Loam (Trevino soila) in Power County, Idaho
|
|
South Exposure |
North Exposure |
||||
|
Major Species |
Production (kg/ha) |
Composition (percent) |
Frequency (percent) |
Production (kg/ha) |
Composition (percent) |
Frequency (percent) |
|
Bluebunch wheatgrass (Agropyron spicatum) |
165* |
27 |
84 |
172* |
24 |
100 |
|
Sandberg bluegrass (Poa secunda) |
99* |
17 |
100 |
119* |
17 |
100 |
|
Thurber needlegrass (Stipa thurberiana) |
77* |
12 |
100 |
62* |
9 |
100 |
|
Balsamroot (Balsamorhiza sagittata) |
43* |
7 |
12 |
78* |
11 |
29 |
|
Hawksbeard (Crepis acuminata) |
39* |
6 |
38 |
53* |
7 |
51 |
|
Longleaf phlox (Phlox longifolia) |
20 |
3 |
83 |
26 |
4 |
88 |
|
MacDougal lomatium (Lomatium macdougali) |
17 |
3 |
62 |
1 |
1 |
1 |
ties. There was a significant difference in total production, however, with the site exposed to the north producing an average of 17 percent more biomass than that produced on the site exposed to the south. The two exposures were classified as different sites (Shiflet, 1973).
ECOLOGICAL SITES AND TYPES
The site classifications proposed for use by BLM for rangeland classification are similar to those used by SCS. They differ primarily in terminology rather than concepts (Society for Range Management, Range Inventory Standardization Committee, 1983). BLM's system is based on ecological sites (BLM) that would be expected to produce a characteristic potential natural community (USFS and BLM) that has a predictable plant composition and annual production. Potential natural community (BLM) describes a plant community composition that accepts some naturalized nonnative species in that community. This differs from the SCS definition of climax plant community (SCS) (see above).
The USFS is changing its classification of rangelands. It uses the term ''ecological type'' (USFS) to classify its rangelands. The term is defined in a newly released USFS manual as ''a category of land having a unique combination of potential natural community, soil, landscape features, climate, and differing from other ecological types in its ability to produce vegetation and respond to management" (U.S. Department of Agriculture, U.S. Forest Service, 1991b:Section 2090.11-05). Descriptions of vegetation (potential natural community [USFS and BLM] and successional stages), soils, topographic features, water, climate, geology, and management interpretations are to be included in ecological type (USFS) descriptions.
The proposed USFS classification system is similar to the SCS system, but it differs in the definition of potential natural community (USFS and BLM). USFS defines potential natural community as follows: "The biotic community that would be established if all successional sequences of its ecosystem were completed without additional human-caused disturbances under present environmental conditions. Grazing by native fauna, and natural disturbances, such as drought, floods, wildfire, insects, and disease, are inherent in the development of potential natural communities which may include naturalized non-native species" (U.S. Department of Agriculture, U.S. Forest Service, 1991b:Section 2090.11-05).
Descriptors of potential natural community (USFS and BLM) include, at a minimum, (1) a list of plant species on the site; (2) some measures of plant species composition or the dominant plant species; (3) production parameters such as weight, cover, basal area (the cross-sectional area of plant stems), incremental growth, or site index (an indicator of site pro-
ductivity based on measuring the height to which a tree on the site has grown in 50 years); (4) a measure of constancy (an indication of the likelihood of finding a species in a given community), by species; and (5) general environmental data. "Ideally, a well described [potential natural community] will show all communities within the sere [a sere is one of a series of ecological communities formed in ecological succession] to be expected following different kinds of disturbance" (U.S. Department of Agriculture, U.S. Forest Service, 1991b:Section 2090.11-2.15).
Evaluation of Range Condition and Ecological Status
SCS, USFS, and BLM evaluate successional change on rangelands by comparing the composition and annual biomass produced by the existing vegetation with a previously determined benchmark plant composition and production. This benchmark is defined by SCS as the climax plant community (SCS) for that range site (SCS), and it is defined by USFS and BLM as the potential natural community (USFS and BLM) for that ecological type (USFS) or ecological site (BLM), respectively. SCS defines the term "range condition" (SCS) as "the present state of vegetation of a range site in relation to the climax plant community for that site" (U.S. Department of Agriculture, Soil Conservation Service, 1976:Section 305.2). USFS defines the term "ecological status" (USFS) as the ''degree of similarity between existing vegetation (all components and characteristics) and soil conditions compared to the Potential Natural Community and the desired soil condition on a site'' (U.S. Department of Agriculture, U.S. Forest Service, 1991b:Section 2090.11-05). BLM also uses the term "ecological status" (USFS and BLM) to describe ''the present state of vegetation in relation to the potential plant community" (U.S. Department of the Interior, Bureau of Land Management, 1985a:Section). All current systems depend on a comparison of the current plant composition with an established benchmark plant composition.
RANGE CONDITION (SCS)
The range condition (SCS) on a given site is based on concepts of succession and retrogression toward or away from the defined climax plant community (SCS). That is, the climax species composition of a particular range site (SCS) changes because of grazing, climatic cycles, fire, insects, physical disturbances, and other stresses (U.S. Department of Agriculture, Soil Conservation Service, 1976). These stresses result in the replacement of species that are characteristic of the climax plant community (SCS) composition by other species characteristic of earlier successional stages. Rangeland managers measure the species composition of a
site and compare it with that expected in the climax plant community (SCS). The similarity is the range condition (SCS). A retrogression (decline) in range condition (SCS) is assumed to be a predictable change from one set of species to another when the same disturbance (for example, grazing or drought) is working. Likewise, the recovery of a site is assumed to be a predictable process of succession. Plants that grow on the deteriorated area are replaced by species that are typical of a later stage of succession until the climax plant community (SCS) is ultimately restored.
Range condition (SCS) classification was originally designed to measure the influence of livestock grazing on the composition and production of the plant community. The hypothesis was as follows:
Grazing management determines place, time, and amount of foliage removal. Removal of green foliage by grazing retards growth most among the species grazed most. This favors the species grazed least because more of the water, nutrients, and light available per unit of surface is left for them. Thus, without practical management of grazing, the normally shorter species are favored as well as the least palatable and the annum species. As the taller species lose ground under close grazing, their place is taken by species short enough to escape with a high percentage of their foliage ungrazed. The result is general reduction in yield as well as measurable change in species composition.
On rangelands, this process, fortunately, can be reversed. Ordinarily a change in management of grazing is all that is required. If secondary plant succession is permitted or fostered, the combination of plants that produces the greatest tonnage of foliage will crowd out other combinations of plants resulting from past mismanagement. Foliage on rangeland in top condition is almost all forage (Dyksterhuis, 1958:151).
The link between grazing pressure and condition class was also thought to be clear and direct.
It follows that the range condition classification provides a measure of determining and stating specifically how much a pasture is overgrazed. In the range-condition system the amount of improvement possible is the exact reciprocal of the amount overgrazed ....
Accordingly, it is more informative to name the condition class than to point out overgrazed areas. Excellent range condition means the site is not overgrazed, while good, fair, and poor conditions refer to three specific grades of past overuse that show today, but can be corrected through one, two, or three classes (Dyksterhuis, 1958:154).
To determine the range condition (SCS) of a site, SCS measures the amount (in kilograms per hectare or pounds per acre) of annual production (air dry weight) by clipping and weighing the biomass or by using standardized procedures to estimate the amount of the current year's plant growth in randomly located plots (U.S. Department of Agriculture,
Soil Conservation Service, 1976). The clipped or estimated weights determined for individual plant species are divided by the total weight of all species to determine the relative percent composition of each species. The percentage of each species is compared with the percentage expected in the climax plant community (SCS) to determine range condition (SCS). Standardized forms are provided by each state SCS office for use in making these calculations and recording the ratings. The forms provide employees with the option of recording other information about the particular site, for example, erosion treatment needs and special considerations such as riparian areas.
Sites occupied by 76 to 100 percent of their climax plant community (SCS) species are rated in excellent condition. Those occupied by 51 to 75 percent of the climax plant community (SCS) species are rated as good, those with by 26 to 50 percent of the climax plant community are rated as fair, and those with 25 percent or less of their climax vegetation are rated as poor (U.S. Department of Agriculture, Soil Conservation Service, 1976).
Although the species composition information described above is the most important factor in determining range condition (SCS), SCS rangeland conservationists can lower the condition rating if certain conditions are present. These conditions include the following:
-
if the overall productivity of the plants on the site is lower than normal and cannot be explained by abnormal conditions (for example, drought);
-
if many of the plant species expected to be found on the site are missing; and
-
if there are signs of accelerated erosion (U.S. Department of Agriculture, Soil Conservation Service, 1976).
Table 3-4 shows how range condition (SCS) can be determined for a particular range site (SCS). The table lists the major species expected in the climax plant community (SCS) characteristic of the Middle Cobbly Loam range site (SCS) in Box Elder County, Utah. Cheatgrass (Brornus tectorum) and snakeweed (Gutierreza sarothrae) are not considered part of the climax plant community (SCS) for the Middle Cobbly Loam range site (SCS), but they were found on the site being rated, so they are listed as having zero potential composition. The potential species composition is the benchmark against which the site to be rated is compared. The estimated percent composition of each species found during the investigation of the site can then be used to calculate the range condition (SCS) rating. This comparison of the actual plant composition with the composition of the climax plant community (SCS) is illustrated in Figure 3-1.
Allowable composition can be equal to, but cannot exceed, the potential percent composition for each species expected in the climax plant
Table 3-4 Determination of a Range Condition (SCS) Rating for Middle Cobbly Loam Range Site (SCS) in Box Elder County, Utah
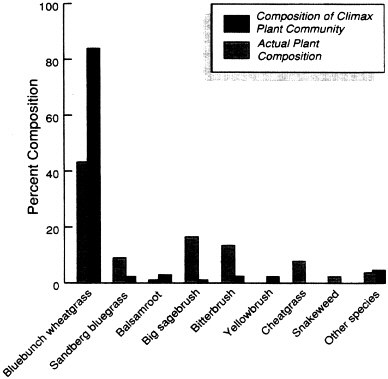
FIGURE 3-1 Comparison of actual plant composition with climax plant community composition for range condition (SCS) rating. Source: Derived from T. N. Shifter. 1973. Range sites and soils in the United States. Pp. 26-33 in Arid Shrublands: Proceedings of the Third Annual Workshop of the United States/Australia Rangeland Panel, D. H. Hyder, ed. Denver: Society for Range Management.
community (SCS). The potential composition of bluebunch wheatgrass (Agropyron spicatum), for example, is 85 percent, and the estimated composition of bluebunch wheatgrass on the site being rated is 45 percent; therefore, 45 percent is recorded as an allowable composition for blue-bunch wheatgrass in Table 3-4. On the other hand, the potential composition of sandberg bluegrass (Poa secunda) is 2 percent, but it was estimated at 9 percent on the rangeland being rated. Since the allowable composition cannot exceed the potential composition, only 2 percent is recorded for the allowable composition of sandberg bluegrass in Table 3-4. Cheat-grass and snakeweed were estimated to be present at 8 and 2 percent compositions, respectively. Neither species, however, is considered part
of the climax plant community (SCS) for the Middle Cobbly Loam range site so both species are given 0 percent allowable percent composition.
This process is followed until all of the species found on the site being rated are assigned an allowable percent composition. The total of the allowable percent composition of each species is then used to determine the range condition (SCS) rating. In the example presented in Table 3-4, the range condition (SCS) rating is 55, meaning that the current composition of the rangeland is 55 percent of its potential climax plant community (SCS) composition. The rangeland is then assigned to a range condition class on the basis of its range condition (SCS) rating as described above. The range condition of the rangeland described in Table 3-4 would be good. The range condition (SCS) class of the Middle Cobbly Loam range site could be lowered from good to fair if the investigator noted and recorded signs of serious erosion or if total production was substantially lower than expected for the Middle Cobbly Loam range site in good range condition (SCS).
The SCS office in each state has developed a form to be used by its personnel when doing a range condition (SCS) rating. (See the Appendix for an example of such a form.) The appearance of the form may differ from state to state, but the procedure is standardized.
ECOLOGICAL STATUS (USFS AND BLM)
The BLM system measures the departure from the potential natural community (USFS and BLM) plant composition and production in much the same way that SCS measures the departure from the climax plant community (SCS) composition (U.S. Department of the Interior, Bureau of Land Management, 1985a). An ecological status (USFS and BLM) rating, which is calculated as a percentage of the potential natural community composition and production, is calculated on the basis of field measurements of plant composition and production by weight. Ecological status (USFS and BLM) ratings are reported in the following four classes:
-
Early seral—0 to 25 percent of the potential natural community (USFS and BLM) is being produced.
-
Midseral—26 to 50 percent of the potential natural community (USFS and BLM) is being produced.
-
Late seral—51 to 75 percent of the potential natural community (USFS and BLM) is being produced.
-
Potential natural community (USFS and BLM)—76 to 100 percent of the potential natural community (USFS and BLM) is being produced (U.S. Department of the Interior, Bureau of Land Management, 1985a).
These ecological status (USFS and BLM) ratings are based on the depar-
ture of the present plant community from the potential natural community. As such, the status is a reflection of current vegetation in relation to the established potential natural community (USFS and BLM) for that ecological site (BLM).
USFS uses ecological status (USFS and BLM) as its measure of the degree of similarity between the existing vegetation and soil conditions compared with those of the potential natural community and the desired soil condition on a site. This similarity can be expressed on a relative scale ranging from 0 to 100, with adjectival ratings assigned as low, moderate, or high similarity. USFS methods of determining ecological status (USFS and BLM) are similar in approach to those of SCS. The frequency of occurrence, however, rather than an estimate of production from clipped weights is sometimes used as the measure of plant composition (U.S. Department of Agriculture, U.S. Forest Service, 1991b).
Trend and Apparent Trend
The concept of trend (a change in a certain characteristic of a rangeland over time) has been used since the early 1900s. It provided a simple, easily applied, and cost-effective method for determining whether grazing practices were causing the desired change in rangelands. Increased soil cover might, for instance, be a goal of management. After the correct stocking rate is calculated, a baseline measure of vegetative cover is made. Cover measurements are made on the same area on a yearly or other established schedule. A downward trend usually results in further adjustments in the level or liming grazing allowed on the site.
Interpretation of the trend concept as a measure of change in range condition (SCS) or ecological status (USFS and BLM) became standardized and accepted in rangeland inventorying and monitoring. A trend in either range condition (SCS) or ecological status (USFS and BLM) is now strictly defined as an evaluation of whether the plant composition and production of a particular rangeland is becoming more similar or less similar to the defined climax plant community (SCS) or potential natural community (SCS) for that particular rangeland site (or ecological type [USFS] or ecological site [BLM]).
Trend is determined by repeated evaluation of range condition (SCS) or ecological status (USFS and BLM) at the same location but at different points in time. Managers, however, often needed a more immediate assessment of trend to evaluate the effect of a change in management. Range scientists attempted to develop a set of criteria that would indicate a trend in ecological state from an evaluation at one point in time. This evaluation of trend has been referred to as apparent trend (SCS).
SCS lists several characteristics of vegetation and soil that can be used
to indicate apparent trend, including the abundance of later-succession (desirable) seedlings and earlier-succession (undesirable) young plants, accumulation of plant residues, plant vigor, and the condition of the soil surface.
According to SCS, the relative importance of these trend indicators varies depending on the site's vegetation, soils, and climate. Any single indicator will give some indication of change in range condition (SCS), but SCS recommends that the evaluation of apparent trend (SCS) be made by considering all indicators in their proper relation to each other (U.S. Department of Agriculture, Soil Conservation Service, 1976).
Apparent trend (SCS) is a professional judgment made at one point in time. It is based on the current status of the vigor, reproduction, distribution, age, and other characteristics of the vegetation as well as on soil, litter cover, erosion, and other factors of a site.
NEW METHODS NEEDED TO ASSESS RANGELAND HEALTH
The current range condition (SCS) or ecological status (USFS and BLM) ratings systems have four components: (1) the classification of rangelands into sites based primarily on differences in their expected climax plant communities (SCS) or potential natural communities (USFS and BLM), (2) the rating of condition or status as the degree to which the current plant composition and production represent those of the benchmark climax plant community (SCS) or potential natural community (USFS and BLM), (3) estimation of site potential on the basis of the plant composition and production estimated for the defined climax plant community (SCS) or potential natural community (USFS and BLM), and (4) estimation of whether a rangeland is changing in ways that will make it more or less similar to the defined climax plant community or potential natural community (USFS and BLM).
The current system of rangeland assessment will not serve as an adequate evaluation of rangeland health, as defined by the committee. The current system does not adequately assess soil stability or the integrity of ecological processes such as nutrient cycles and energy flow. There are problems with each of the four components of the current system that will limit its utility as a measure of rangeland health.
Site Classification
Even the casual observer can look at a rangeland and recognize that some parts of the landscape are different from others. These differences are expressed in many ways, but they are expressed most obviously
through topography, vegetation, and soil surface characteristics like rocks, bare soil, or litter cover.
In practice, the resulting site delineations are arbitrary landscape divisions that can be used to provide order to a complex system for management purposes and extrapolation of research results (Passey and Hugie, 1962; Pendelton, 1989; Tueller, 1973). The major use of the site concept in rangeland assessment has been for analysis of range condition (SCS) or ecological status (USFS and BLM). Sites therefore have most often been delineated by studying the composition and production of climax community plant species (Shiflet, 1973).
CLASSIFICATION BASED ON CLIMAX CONCEPT
A focus on climax plant community (SCS) or potential natural community (USFS and BLM) composition for distinguishing between sites is the common point of departure for all site classification systems used by federal agencies. Differences in the expected plant composition and production in the climax plant community have been used as a way of distinguishing the variability in plant composition caused by use, management, stage of succession, or other variables from the variability caused by differences in the site.
The reliance on differences in the expected climax plant community (SCS) or potential natural community (USFS and BLM) composition for distinguishing between sites, however, has been questioned on both theoretical and practical grounds. Plant composition at any one point in time varies because plant communities are constantly changing in composition and production owing to changes in environmental influences (Gleason, 1926; Harper, 1977). Some scientists have questioned whether the concept of a single, definable, and predictable climax plant community can be applied to all rangelands (West, 1985; Wilson, 1989). Others have suggested that succession may follow multiple pathways and that the pathway followed by a particular rangeland depends on the kind of disturbance and the environmental conditions during secondary succession (Friedel, 1991; Risser, 1989; Smith, 1989; Westoby et al., 1989). The plant community that arises from primary succession (succession that begins on a habitat that has not previously been inhabited), therefore, may be different from the plant community that arises from secondary succession (succession that follows destruction of all, or part, of a previous plant community). Differences in expected successional processes and their resulting end points may be due as much to the type of disturbance and the prevailing conditions at the time of disturbance as to differences between sites.
The practical difficulties of determining climax vegetation have also been cited. Although there may not be relic (undisturbed) rangelands that
can be used as benchmarks of climax plant community composition (Shiflet, 1973), several methods can be used to reconstruct a hypothetical climax plant community: (1) interpolation and extrapolation from existing relic areas, (2) comparison of grazed with ungrazed areas, (3) evaluation and interpretation of research data on plant communities, and (4) review of historical accounts and botanical literature (Pendelton, 1989). Such a process can be a difficult and time-consuming endeavor, however.
Alternative Approaches to Site Classification
Because of the problems with basing site distinctions on differences in climax vegetation, several general suggestions for change have been made by various groups and individuals.
Bentley and Talbot (1951) classified different kinds of rangelands on the basis of the amount of vegetation each type of rangeland could produce, without regard to species, on California grasslands where exotic grasses that have been introduced by humans dominate. On these sites, estimation of the botanical composition of the climax vegetation has little relevance to current management since the ability to return to climax vegetation has been eliminated by the introduction of exotic species.
The Range Inventory Standardization Committee of the Society for Range Management (1983) recommended the term "ecological site" for the basic unit of rangeland classification. That committee defined ecological site as a kind of land that differs from other kinds of land in its potential natural community and physical site characteristics and, therefore, also differs in its ability to produce vegetation and its response to management.
Recently, the Society for Range Management recommended that land types that differ significantly in their ability to produce vegetation (either kind or amount) should be called ecological sites and should be defined as a kind of land with physical characteristics that differ from those of other kinds of land in their ability to produce distinctive kinds and amounts of vegetation and in their response to management. The difference in this definition from that of the earlier committee (Society for Range Management, Range Inventory Standardization Committee, 1983) is the omission of a reference to a potential natural community (USFS and BLM). Justification for using ecological site (BLM) rather than range site (SCS) is based on the reasoning that site classification is not necessarily oriented to any particular land use or land type.
Considerable evidence exists in the ecological and agronomic literature for the classification of rangelands into sites on the basis of relationships among climate, soil, and vegetation. In nature, these boundaries are arbitrary, meaning that all site descriptions are approximate but useful
classifications that can be used to organize management knowledge and research results.
Because site classification is an arbitrary division of the landscape and because individual sites exist within a patchwork of other sites, some considerations of influences at the landscape level may be desirable for some purposes. Australian researchers have developed land systems models that describe sites that occur together on rangelands (Mabbut, 1968). Similar to soil associations, these land systems models relate adjacent sites that may or may not have common characteristics. Analysis of the ways in which sites are associated and their interactions within a landscape could improve understanding of ecosystem processes at the multiple-site level, where most management implementation occurs. For example, landscape position may be important when sites that receive runoff, such as riparian zones, are being eroded because of vegetation conditions on other parts of the watershed.
SITE CLASSIFICATION FOR ASSESSING RANGELAND HEALTH
SCS, USFS, and BLM should adopt common site classifications for the purpose of coordinating rangeland health inventories and monitoring efforts.
Although methods of site classification between the agencies are similar in concept, differences in the definitions of climax plant communities (SCS) and potential natural communities (USFS and BLM) make comparisons of assessments on different administrative units difficult. It is important that all three agencies (BLM, SCS, and USFS) use similar classification methods, so that the results of management, research, and assessments can be compared across administrative boundaries.
To limit interpretation conflicts that could arise from the proliferation of various classification schemes for various purposes, common site classifications should be soil based and should provide general information on vegetation production and life-form dynamics. They should also describe responses to disturbances such as fire, grazing, and drought. For the purposes of rangeland health assessments, it would be useful to add descriptors of soil surface, nutrient cycles, energy flows, and recovery mechanism attributes to the current criteria for describing range sites (SCS) or ecological types (USFS).
Range Condition: Ecological Status
Traditionally, range scientists have defined the term ''range condition'' to mean ''the state of range health" (Society for Range Management, 1989:2), and federal management agencies have, in fact, used the term in this manner (see, for example, U.S. Department of the Interior, Bureau of
Land Management [1984], and U.S. Department of Agriculture, Soil Conservation Service [1989a]). The development of the current methods for evaluating the ecological state of rangelands on the basis of the departure from climax vegetation and the succession-retrogression model of rangeland change can be viewed as the first approximation of rangeland health. There were and are reasons to consider rangelands that contain climax vegetation healthy, as defined by the committee. The process of ecosystem development—that is, succession—was thought to culminate in maximum stability, productivity, diversity, and other presumed desirable qualifies (Stoddart et al., 1975). Communities in the early stages of succession were thought to be characterized by less complex energy flows and more open nutrient cycles and to be more vulnerable to invasion by exotic species (Odum, 1969). Assessments of range condition (SCS) and ecological status (USFS and BLM) have produced a wealth of useful data and research that has provided the underpinning for efforts to manage the impact of grazing on both federal and nonfederal rangelands.
Range condition (SCS) and ecological status (USFS and BLM) ratings, however, are not sufficient measures of rangeland health, as defined in this report. The committee identified three problems with range condition (SCS) and ecological status (USFS and BLM) that limit the utility of these methods as measures of rangeland health: (1) use of climax plant community (SCS) or potential natural community (USFS and BLM) composition as standards, (2) the difficulty in identifying thresholds of change, and (3) the reliance on changes in plant composition and production as the sole indicator of change in the ecological state of rangelands.
STANDARDS FOR ACCEPTABLE CONDITIONS
The current methods of assessing range condition (SCS) or ecological status (USFS and BLM) establish a benchmark plant community against which current plant composition and production are compared. Condition and status ratings are a measure of how closely the current vegetation resembles the defined benchmark plant community.
LINKING SUCCESSIONAL STAGE TO CONDITION AND STATUS RATINGS The linking of successional stages to range condition (SCS) classes has confused the interpretation of the results of range condition (SCS) and ecological status (USFS and BLM) surveys. Smith (1989) has observed that acceptance of the view that succession is ecosystem development that culminates in maximum stability, productivity, diversity, and other presumed desirable qualities "really leaves one with little choice but to manage for near climax, or admit that the goal is a second rate, degenerated ecosystem" (Smith, 1986:120). The public, the U.S. Congress, and environmental in-
terests are understandably concerned with reports that 6 and 16 percent of lands managed by BLM are in fair or poor condition, respectively. Concern that poor or fair condition does indicate rangeland degradation is reinforced when decreases in the amount of rangelands in poor or fair condition are reported as evidence of agency success in meeting mandates to improve rangelands (see U.S. Department of the Interior, Bureau of Land Management [1990], for example). Rangeland managers and livestock producers respond that fair and poor conditions do not necessarily indicate a problem or the need for changes in management practices on a particular site; they only indicate that rangeland's stage of succession. BLM and USFS have eliminated the terms excellent, good, fair, and poor and have adopted terminology that reflects successional stages. The problem that remains, however, is in determining whether there is a cause for concern about any one of the successional stages that a rangeland may be in.
The relationship between successional stages and the stability of a site's soil and the integrity of its ecological processes—that is, its health—is uncertain. Spence (1938) noted that the soil, water, and productiveness of a rangeland are conserved when it contains its climax vegetation but that rangelands in earlier stages of succession can also conserve these values. Spence also noted that species that are not part of the climax vegetation can also conserve the soil, water, and productiveness of rangelands.
Lauenroth (1985) concluded that the Similarity of a rangelands' vegetation to climax vegetation does not necessarily indicate where site degradation is occurring or where it might occur. Different combinations of plants with the same degree of similarity to an established benchmark community may differ in the effectiveness with which they protect the site from accelerated soil erosion. Risser (1989) noted that a rangeland's vegetative composition may not reflect the erosional status of the soil mantle, and Wilson (1989) reported that there is no clear relationship between changes in plant composition and soil erosion on rangelands studied in Australia. Smith (1989) also noted climax vegetation is not the only type of vegetation that furnishes adequate soil protection. Other investigators have noted that seedlings of exotic species, such as crested wheatgrass (Agropyron cristatum), that are completely dissimilar to the climax vegetation can offer adequate protection from erosion (Dormaar et al., 1978).
The committee inspected a site near Reno, Nevada, that illustrates the difficulty in clearly relating the degree of similarity to an established benchmark plant composition to the degree to which the soil and ecological processes are being conserved. The existing plant composition was dominated by desert needlegrass (Stipa speciosa) and Wyoming big sagebrush (Arteraisia tridentata ssp. wyomingensis ) but included a number of
other species such as Indian rice grass (Oryzopsis hymenoides) and Utah juniper (Juniperus osteosperma). The percent species composition was similar enough to the climax plant community (SCS) so that the rangeland could be rated in the excellent condition class on the basis of the rangeland's plant composition alone. The range condition (SCS) of the site was reduced to good, however, because of serious wind erosion. Wind erosion on the site was severe enough that many plants were being damaged by abrasion, and some were being buried by drifting soil. It appeared that the amount of plant cover on the site had declined, exposing the soil to wind erosion. The decline in plant cover, however, appeared to have been evenly distributed among all species present on the site, so that the percent composition remained the same as that which would be expected in the climax plant community (SCS). Similarity to a benchmark plant composition was not, in this case, a sensitive indicator of other changes that were occurring on the site.
The similarity of current plant composition and biomass production to that of a climax plant community (SCS) or potential natural community (USFS and BLM) should not be used as the primary standard of rangeland health.
The degree of similarity to the plant composition and annual biomass production of a climax plant community (SCS) or potential natural community (USFS and BLM) alone is not a sufficient measure of rangeland health. The evaluation of rangeland health will require additional and different criteria and indicators.
DIFFICULTIES IN COMPARING DIFFERENT SITES The climax plant community (SCS) and the potential natural community (USFS and BLM) is different for each site. Similarly, the plant composition and biomass production of successional stages also may differ between sites. Since the benchmark against which current plant composition is compared is different for different sites, comparing the results of condition or status ratings between sites is difficult. It is possible, for example, that an early successional stage on one site may closely resemble a later successional stage on a different site. Plant composition and production may be very similar between the two plant communities on the two sites, but they may receive different condition or status ratings since the benchmarks against which they are being compared differ.
Laycock (1989) cited an example of such a problem in Idaho:
The Artemisia tridentata ssp. wyorningensis/Poa secunda [Wyoming big sagebrush/sandberg bluegrass] habitat type occurs in areas in western Idaho where precipitation is less than 18 cm annually (Hironaka et al., 1983). The dominant plant is the Wyoming big sagebrush with an understory of sandberg bluegrass and scattered other species. This [Wyoming
big sagebrush/sandberg bluegrass] vegetation mix is identical to a rather severely degraded ("poor condition") stage of the Artemisia tridentata wyomingensis/Stipa thurberiana [Wyoming big sagebrush/Thurbers needlegrass habitat] type that occurs over a wide area of southern Idaho on better sites and higher precipitation where grazing has removed the [needlegrass] and other desirable species. It may also resemble other sagebrush habitat types in lower stages of condition. (Laycock, 1989:5)
The sites described by Laycock had essentially the same plant compositions and production, but they would have different ratings since the benchmarks against which they were compared differed.
This difference may be important because the relationship between a similarity rating and the conservation of soft and ecological function is not well understood. One site that supports 40 percent of its expected climax plant community (SCS) or potential natural community (USFS and BLM) composition may still have enough soil cover to be protected from water and wind erosion, whereas another site, also supporting 40 percent of its expected climax plant community (SCS) or potential natural community (USFS and BLM) composition, might be eroding at rates that will lead to serious site degradation, it might have a compacted soil that restricts infiltration, or it might have nutrient cycles that are interrupted by a lack of litter cover and lack of incorporation of organic matter into the soil surface.
The use of similarity to a defined benchmark plant community, whether defined as climax plant community (SCS) vegetation or potential natural community (USFS and BLM), also imposes the difficulty of direct comparisons between sites with different benchmark plant communities. The lack of a single, dearly defined standard that does not differ from site to site is a fundamental limitation to the use of current methods for assessing rangeland health. This problem has and continues to confuse the public, the U.S. Congress, livestock producers, and rangeland scientists themselves.
LIMITATIONS OF SUCCESSION-RETROGRESSION MODEL
The successional model that supports the current range condition (SCS) and ecological status (USFS and BLM) methods postulates that succession toward the climax plant community (SCS) or potential natural community (USFS and BLM) is a process that moves through recognizable and predictable stages. The effects of grazing, drought, and other disturbances are also thought to produce recognizable and predictable changes in composition toward early stages in succession. The vegetation that is found on any particular rangeland is the product of the equilibrium between the two forces of succession toward the climax plant community and retrogression toward earlier successional stages.
This rangeland succession model assumes that range condition (SCS)—that is, the successional stage of a particular rangeland—will change back and forth along the successional gradient characteristic of that site in response to changes in management. The main tool of rangeland managers is to adjust the stocking rate, species of grazing animal, the duration of grazing, and the season that the rangeland is used to achieve an equilibrium between the opposing forces of succession and retrogression. Once this equilibrium is achieved, it will tend to remain stable. The manipulation of livestock grazing allows rangeland managers to maintain a particular plant composition at some point along the successional gradient characteristic of that site. The choice of that point along that gradient that should be maintained depends on whether the goal of management is to maximize or optimize the production of livestock, wildlife, or some other product or value.
DOES SUCCESSION OCCUR ON ALL RANGELANDS? Current ecological research has questioned whether the concept of well-defined, predictable, and reversible changes along a successional gradient holds for all or the majority of rangelands.
Ecologists working in rangeland ecosystems have developed theories that allow for multiple equilibria and for transitions between alternative vegetational states that are not easily reversible (Friedel, 1991; Westoby et al., 1989). These theories, which were discussed in more detail in Chapter 2, also allow for the existence of transitional states that represent the process of change from one state of equilibrium to another. The specific outcome of the change will depend on events that occur while the rangeland is in that transitional state rather than on a predictable succession from one state to another. Investigators have attempted to describe the mechanisms that produce such complex dynamics on rangelands. In some cases, the random occurrence of fire, drought, or changes in grazing systems have produced changes in rangelands that do not appear to follow a readily discernible successional sequence (Friedel, 1991; Laycock, 1989; Westoby et al., 1989).
Sharp et al. (1990), for example, reported 40 years of data from a salt desert shrub rangeland in south-central Idaho that illustrate this phenomenon. Three different plant communities have occurred on this site, which has not been grazed since 1945. During periods of normal precipitation and no buildup of a naturally occurring scale insect (Orthisae sp.), the site is dominated by shadscale (Atriplex confertifolia) in association with bottlebrush squirreltail (Sitanion hystrix), sandberg bluegrass (Poa secunda), globe mallow (Sphaeralcea coccinea), and other plants. Following a cyclic outbreak of the scale insect, most of the shadscale dies or is reduced in size and vigor. If precipitation is normal in the years following the out-
break, the site becomes dominated by bottlebrush squirreltail, production is high, and shadscale may eventually recover. If precipitation is below average following the insect outbreak, several species codominate the site. The species that becomes more important depends on the timing of precipitation events. The transition from one complex to another appears to be due more to the vegetation's adaptation to episodic events than to a linear successional development. It could be argued, however, that the desert sites described simply have not had time to recover from historic overgrazing and to reestablish the potential natural community (USFS and BLM).
In any case, the kinds of vegetation dynamics described above are difficult to incorporate into existing succession-retrogression models of rangeland development.
Risser (1989) summarized the questions about the succession-retrogression model raised by ecologists as including some of the following ideas. Biological communities may go through different pathways yet reach a similar climax or terminal state. Depending on disturbances during succession, the system may proceed through a new set of seral stages. The initial conditions at the beginning of a successional sequence can cause quite different outcomes even on apparently equivalent sites. The outcome of successional sequences is determined by the characteristics of the interacting plant and animal populations and the present and preceding environment, not by predetermined organismic controls. Certain ecosystem-level characteristics, such as the ability to absorb or release nutrients, are characteristic functions of systems in early and late stages of succession. The terminal stage may not be the most productive, stable, or diverse community.
Risser concluded that it is important to recognize that a simple linear successional sequence is not always an adequate representation of the conditions observed in the field (Connell and Slatyer, 1977; Drury and Nisbet, 1973; Gutierrez and Fey, 1975; McIntosh, 1980; Odum, 1969 [as cited by Pisser, 1989]).
Traditional successional theory implies that a site that has retrogressed can recover if the process is reversed. This is not possible or is very slow, however, if severe soil erosion, invasion of a new and very dominant species, or change from a fire-dependent to a fire-safe plant community has resulted in near-permanent changes in the abiotic or biotic community.
The succession-retrogression model of how rangelands develop and change, which is the foundation of current rangeland classification and assessment methods, should be modified or new models developed to assist in assessing whether rangelands are approaching thresholds of change.
Current range condition (SCS) and ecological status (USFS and BLM) ratings are founded on the concept of well-defined, predictable, and reversible changes along a successional gradient that holds for all or the majority of rangelands. An evaluation of rangeland health requires consideration of additional processes of ecosystem change and the reversibility of those changes. Several alternative models of rangeland change have been proposed, but no single model has received widespread acceptance. An accelerated effort by ecologists is needed to develop and test models of rangeland change that will assist in identifying rangelands that are approaching thresholds of change.
MULTIPLE INDICATORS ARE NEEDED
Current range condition (SCS) or ecological status (USFS and BLM) ratings rely nearly exclusively on measurements of plant composition and annual biomass production. Problems such as soil erosion, disruption of nutrient cycling, or other ecological attributes of rangelands are not primary considerations in assessing range condition (SCS) or ecological status (USFS and BLM). Changes in other important attributes of an ecosystem may not be detected by measuring the plant composition and production alone.
The difficulty of assessing ecosystems using only one index has long been recognized. Ellison (1949) wrote that the soft, plants, animals, topography, and climate develop together and are knit together into an integrated whole. He also noted that vegetative and soft trends do not always parallel each other and that they may be widely divergent on eroding rangelands. He suggested that "if the rangeland manager is impressed by evidence of change in vegetation and not by evidence of soil erosion, he may be led astray" (Ellison, 1949:794).
The effectiveness of vegetation in protecting soil is more a function of effective soil cover than plant composition, since effective soft cover is more closely tied to the type and pattern of cover than it is to plant composition. Slight changes in plant populations and litter may produce accelerated erosion on steep, erodible soils, whereas complete changes in species and life-forms, such as in artificially established annual grasslands, may result in a vegetation type that still provides good soil protection. Erosion hazard on some soils may be quite insensitive to changes in vegetation type if the site is very flat or very rocky, whereas on steep day slopes the kind or amount of vegetation may be critical (Smith, 1989).
Loss of minor species may not be indicated by a change in range condition (SCS) or ecological status (USFS and BLM) rating if these species make up a small percentage of the plant composition and annual biomass production of the climax plant community (SCS) or potential
natural community (USFS and BLM). The loss of minor species, however, may indicate change in nutrient cycles caused by reduced diversity in rooting depth or changes in energy flow because of a reduced period during which the remaining plants photosynthesize.
Tueller (1973) described a process of site degradation that began with the loss of plant vigor and seed production and that led to the death of individual plants and a reduction in litter cover and plant density. These changes caused changes in plant cover, distribution, and potential for reproduction. Total biomass production or the annual production of individual species was reduced. Further deterioration led to reduced litter accumulation, the formation of soil crusts that retarded germination, and altered plant growth forms. Reduction in soil cover and litter led to soil erosion and the disruption of nutrient cycles. Eventually, the site potential was seriously impaired.
The process of site degradation described by Tueller (1973) is driven by a complex of interacting factors, with no single factor predominating. Changes in species composition, plant density or frequency, distribution and cover of litter, soil erosion, total biomass production, plant vigor, and seedling recruitment, among other factors, are all apparent at different points in the degradation process. No single factor alone can completely describe such a process. Tueller for instance, listed 16 separate factors that can serve as useful indicators of site degradation (Tueller, 1973).
The problem is not that plant composition and biomass production are unimportant attributes of rangeland ecosystems; rather, the problem is that they are typically the only attributes measured. A system that used only plant density, erosion, litter cover, seedling density, or compaction as a single measure of range condition (SCS) or ecological status (USFS and BLM) would have the same problem.
The evaluation of rangeland health will require analysis of attributes in addition to plant composition. A comprehensive evaluation of rangelands should be based on the preponderance of evidence derived from sampling multiple attributes related to ecological function and soil stability. Range condition (SCS) or ecological status (USFS and BLM) ratings based primarily on changes in plant composition and annual biomass production alone are not sufficient measures of rangeland health.
Plant composition and biomass production will usually change if soil erosion accelerates, the soil continues to compact, or seedlings fail to become established. However, there may be significant lags between the onset of rangeland degradation and a change in any single indicator of rangeland health. Any single measure will have different sensitivities to different stresses. The measurement of multiple attributes increases the probability that rangeland degradation will be detected early enough for corrective measures to be taken.
Site Potential and Resource Values
In current theory and practice, the rangeland site classification and the definition of site potential are nearly synonymous. The kinds and amounts of vegetation produced in the climax plant community (SCS) or potential natural community (USFS and BLM) are considered the best measure of the potential productivity of a site. Overgrazing, accelerated erosion, or other influences that result in loss of the capacity to produce the plant composition and annual biomass production characteristic of the climax plant community (SCS) or potential natural community (USFS and BLM) are thought to have caused a loss in site potential.
DESIRED PLANT COMPOSITION
Many observers have argued that the plant composition and production desired on a site should depend on how the site is to be used. In its National Range Handbook (U.S. Department of Agriculture, Soil Conservation Service, 1976), SCS states that although the climax plant community (SCS) describes the site potential of a particular site, the goal of management is not necessarily always to achieve climax plant community (SCS) composition and production. Other seral (successional) stages may be better for particular uses. Pisser (1989) stated that the climax vegetation for a given site may not by the most productive or desirable type of vegetation for livestock forage production and that climax vegetation may not be the most appropriate goal of rangeland management if the management objectives include multiple uses or values. Such multiple uses or values may include wildlife, water quality or quantity, recreation, and livestock grazing.
RESOURCE VALUE RATING
Wilson (1989) suggested the need to first define the land use objectives for rangelands. This should be followed by a description of the vegetative structure that will maximize those objectives. The Society for Range Management's Range Inventory Standardization Committee similarly recommended that a system of resource value ratings be used as a measure of the value of vegetation or other features for a particular use (Society for Range Management, Range Inventory Standardization Committee, 1983). The resource value rating would be based on the particular species present, growth forms and foliage types, or other criteria. Each use may have a separate resource value rating. The resource value rating was to be a measure of the suitability or usefulness of the vegetation of an ecological site (BLM) for a specific use.
Most recently, the Society for Range Management's Task Group on Unity in Concepts and Terminology recommended that management objectives should be defined in terms of a desired plant community for each ecological site (BLM). The desired plant community should be defined as follows: ''of the several plant communities that may occupy a site, the one that has been identified through a management plan to best meet the plan's objectives for the site'' (Society for Range Management, Task Group on Unity in Concepts and Terminology, 1991:10).
USFS has proposed adoption of a system of resource value ratings. Resource value ratings require that goals be set for the vegetation needed to meet a particular use. Those goals will be a desired plant community. Because goals will vary from location to location, different desired plant communities may be described for different locations on the same site. Management would be designed so that the plant composition could be changed to reflect that described for the desired plant community. This may or may not represent a change toward the climax plant community (SCS) or potential natural community (USFS and BLM).
DISTINCTION BETWEEN RANGELAND HEALTH, SITE POTENTIAL, AND RESOURCE VALUES
Rangeland health assessments should be separate and independent of assessments for determining the proper use of particular rangelands .
It is essential that there be a clear understanding of the distinction between rangeland health, as defined in this report, and site potential or resource value ratings. Rangeland health, as used in this report, is intended as a minimum ecological standard, independent of the rangeland's use and how it is managed. The particular mix of commodities and values produced by a rangeland depends on how it is used and managed. If rangeland health is conserved, then the capacity of the site to produce different mixes of commodities and values is conserved. The determination of which uses and management practices are appropriate may require the evaluation of data different from those used to evaluate rangeland health. No single index will meet all the needs of rangeland inventory, classification, and management.
Rangeland health is a measure of the integrity of the soil and the ecological processes of a rangeland. Loss of rangeland health causes a loss of capacity to produce resources and satisfy values. Rangeland health is not, however, an estimate of the kinds or amounts of resources that a rangeland produces, nor is it an evaluation of the different uses of a site. Two rangelands may have the potential to produce different commodities and values, but both can be equally healthy if the integrity of the soil and ecological function are conserved.
The protection of rangeland health should serve as the minimum standard for management. If rangeland health is sustained then decisions about the appropriate plant community composition and production can be made depending on the desired rangeland use. Most important, the conservation of rangeland health preserves the option to change the use and management of a site as the desired resources and values change.
Apparent Trend
A one-time measure of most rangeland characteristics is only that—a picture of the situation at the time of measurement. Without a previous measurement with which the current measurement can be compared, the range manager's ability to interpret whether the management program is succeeding or failing is limited. Personnel and budget constraints and inconsistencies in the indicators measured at different times, however, have limited the number of sites where trend can be determined from comparable data collected at different points in time.
To compensate for this problem, range managers have attempted to determine apparent trend; that is, they evaluate site characteristics that indicate whether an area is improving or deteriorating. Factors such as accelerated erosion, for example, have been used to indicate a downward trend. If the soil is eroding at an accelerated rate, then the productive capacity of the site is probably being lost. The accumulation of litter is viewed as a sign of an improving rangeland because it is a sign that the amount of plant material needed to protect a site from erosion is increasing. The presence of seedlings of desirable plants was also interpreted as an upward apparent trend because those seedlings indicated that the plant composition was evolving toward the climax plant community (SCS) or potential natural community (USFS and BLM). The presence of seedlings of undesirable plants represents a downward trend. Plant vigor has also been used to judge apparent trend. Vigorous dominant plants in a climax plant community (SCS) or potential natural community (USFS and BLM) indicate an upward apparent trend, whereas the presence of weak, deformed plants indicates a downward trend.














































