
Dengue: The Risk to Developed and Developing Countries
THOMAS P. MONATH
In the last 20 years, dengue fever and a severe form of the disease described for the first time in the mid-1950s—dengue hemorrhagic fever (DHF)/dengue shock syndrome (DSS)—have emerged as the most important arthropod-borne viral diseases of humans (1–3). During this period the frequency of dengue fever epidemics has increased dramatically, hyperendemic transmission has been established over a geographically expanding range (Figure 1), and DHF has occurred in new areas and at higher incidence. It is estimated that up to 100 million cases of dengue fever occur annually on a worldwide basis. Approximately 250,000 cases of DHF are officially notified, and the true incidence is undoubtedly several-fold higher. Relatively few cases were recognized in the first 25 years after the initial description of this disease, while reports have risen steeply in recent years (Figure 2). The geographic expansion of epidemic dengue fever during the 1980s involved South, Central, and North America; Africa; China; and Australia, and is expected to continue in receptive regions infested by Aedes aegypti. Of greatest concern to the future health of the Western Hemisphere is the emergence of DHF in the Americas during the last 12 years. As illustrated below, the factors responsible for the worldwide increase in the incidence and distribution of dengue are closely linked to changes in human ecology and behavior.
Thomas P. Monath is vice president of research and medical affairs at OraVax, Inc., Cambridge, Massachusetts.
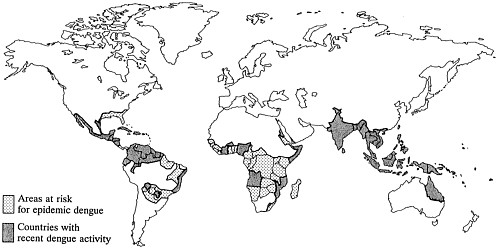
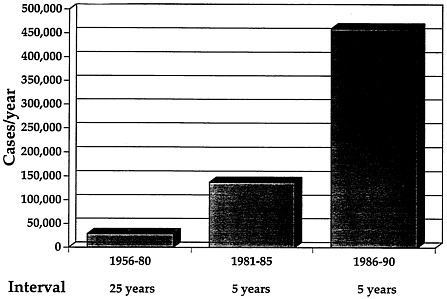
FIGURE 2 Incidence of DHF since its first description as a nosologic entity in 1954(modified from ref. 4). A dramatic increase in incidence has occurred during the 1980s, reflecting both a real increase and an apparent increase due to improved reporting.
TRANSMISSION CYCLE, DISEASE, AND PATHOGENESIS
Dengue is caused by four antigenically distinct single-strand positive-polarity RNA viruses, designated dengue types 1–4, belonging to the family Flaviviridae (5). Virus transmission in its simplest form involves the ingestion of viremic blood by mosquitoes and passage to a second susceptible human host. An extrinsic incubation period of 8–10 days is required after feeding on a viremic human for viral replication and internal dissemination in the mosquito before virus appears in the saliva and transmission on refeeding can occur. As the blood meal stimulates oviposition by the female mosquito, which undergoes at least one, and often more, reproductive cycles during the extrinsic incubation period, there is an opportunity for virus to enter the egg and be passed to the next generation of mosquitoes.
In tropical Asia and West Africa, dengue viruses are also transmitted between nonhuman primates and tree-hole-breeding mosquitoes (6), but it is uncertain what relationship, if any, exists between the forest cycle and the circulation of virus between humans and A. aegypti. However, the existence of a completely silent zoonotic transmission
cycle affords a potential mechanism for emergence of the disease in human populations and possibly also for selection of virus variants with altered host range and vector relationships. Virus strains representing the forest cycle have been subjected to molecular analysis and found to be distinct (see below), indicating that the forest cycle may be ecologically isolated. An important question for future research is whether the virus strains that circulate in the forest cycle are biologically distinct. Nonhuman primates challenged with strains of dengue virus isolated from humans generally develop abbreviated and significantly lower viremias than humans (7). These virus strains have not been evaluated for their capacity to induce viremic responses in their natural monkey hosts. It is possible that the apparent separation of forest and human transmission cycles reflects a reciprocal and exclusive adaptation to their hosts (or vectors).
The uncomplicated disease, classical dengue fever, is a biphasic illness beginning abruptly 3–8 days after the bite of an infected mosquito, characterized by fever, headache, severe malaise, lumbosacral aching, and generalized muscle, joint, or bone pain. Improvement after several days is followed by the reappearance of fever and development of a measles-like rash, generalized lymphadenopathy, and, sometimes, minor hemorrhagic phenomena. There are no fatalities and the disease resolves in the second week, although patients may experience prolonged convalescence, with weakness and depression. Due to the self-limited nature of the infection, little is known about the pathogenesis of classical dengue fever. High titers of virus are present in the blood during the early phase, providing the means for mosquito infection. Dengue virus is predominantly a lymphotropic agent, and the principal target cells for virus replication appear to be mononuclear phagocytes, a fact that assumes greatest relevance in the pathogenesis of DHF/DSS (8).
The onset and early phase of DHF/DSS is identical to that of dengue fever. However, shortly after onset, the patient rapidly deteriorates, developing epigastric pain, restlessness and irritability, thrombocytopenia, and signs of diffuse capillary leakage, hemoconcentration, and hypotension. Hemorrhagic manifestations of all kinds occur. In its most severe form (designated DSS, occurring in up to 1/3 of individuals with DHF), patients experience narrowing of the pulse pressure and circulatory failure. The case-fatality rate of DHF/DSS is up to 20% if untreated, but with supportive treatment consisting of fluid and electrolyte management and oxygen, fewer than 1% of such cases prove to be lethal.
The pathogenesis of DHF/DSS is only partially understood. The disease is an immunopathologic process, dependent in the vast majority
of cases on prior immune sensitization by a heterotypic dengue infection. Infection with one dengue serotype provides lifelong homologous immunity, but only transient cross-protection against other serotypes, making sequential infection possible. The relative risk of experiencing the most severe form of the disease is 100-fold higher after secondary than after a primary infection. The underlying mechanism involves enhanced infection of Fc-receptor-bearing monocyte/macrophages by dengue viruses complexed to nonneutralizing IgG antibodies (8, 9). These antibodies are the result of prior infection with a heterologous dengue virus serotype, or, in the case of infants born to immune mothers, the result of waning passive maternal antibody. The infectious immune complexes gain access to Fc-receptor-bearing monocytes more readily than dengue virus alone, with the result that the host has a larger number of infected cells containing quantitatively higher amounts of dengue virus, a phenomenon known as immune enhancement. A second aspect of the pathogenesis is the marked T-cell activation and induction of cross-reactive CD4+ and CD8+ cytotoxic T cells that recognize dengue viral antigens (principally nonstructural proteins) on infected monocytes (10). Although this process is key to the clearance of infected cells and recovery of the host from infection, the result of this interaction in a subset of patients may also have pathophysiological consequences due to release of cytokines with vasoactive or procoagulant properties (interleukins, tumor necrosis factor, platelet-activating factor, and urokinase), complement activation, and release of interferon γ. The latter molecule up-regulates expression of Fc receptors and in turn increases antibody-dependent enhancement of dengue virus replication. It is still uncertain what host- and virus-specified factors determine why one individual develops DHF/DSS and another clears secondary infection without consequence. Moreover, the precise role of different cytokine mediators in the pathogenesis of DHF/DSS remains to be defined.
HISTORY OF EPIDEMICS AND THE EMERGENCE OF DHF/DSS
Dengue fever has been recognized clinically for over 200 years. During the 18th and 19th centuries the disease occurred in intermittent pandemics affecting Asia and the Americas, occurring at intervals of up to several decades (11, 12). Spread was slow, generally by ships carrying breeding populations of A. aegypti and susceptible human hosts. In many areas, dengue was recognized only among expatriate settlers or colonial military forces, and the disease escaped attention in the indigenous populations under poor medical surveillance.
This pattern changed dramatically during and after World War II. Dengue viruses were spread by viremic military personnel to staging
areas in the Pacific. Multiple dengue serotypes were geographically shuttled by viremic troops and refugees, and the vector was spread by vehicles, water storage containers, and tires carrying along the ova and larvae of A. aegypti. The dissemination of virus and vector was enhanced after the war by rapid population growth and urbanization. Asian cities were characterized by poor sanitation, the necessity for domestic water storage, and crowded living conditions, creating conditions favoring breeding of A. aegypti. Superimposed on these phenomena was the rapid rise in air travel, providing the means for movement of viremic human beings within the region and beyond. These factors led to the establishment of hyperendemic dengue infection in Southeast Asia, a pattern of annual outbreaks caused by all four dengue serotypes, and an increasing frequency of sequential infections of children (13). It is in this setting that DHF/DSS emerged in 1954 in the Philippines. Over the next 20 years, outbreaks occurred that involved many parts of Asia and the Pacific, with a mean annual incidence of about 30,000 cases. In the 1970s and 1980s, the incidence of DHF rose dramatically, to over 250,000 cases per year (Figure 2). DHF is now the third or fourth leading cause of hospitalization of children in some areas (14).
The emergence of dengue fever and DHF in the American region provides a paradigm for the changing features of dengue epidemiology. Prior to World War II infrequent epidemics occurred at intervals of up to 37 years, probably caused by introduction of a single serotype (12). The opportunities for introduction of new viruses were limited. Outbreaks were rarely sustained for more than a few years because human populations were relatively low and isolated in island situations where immunologically susceptible hosts were rapidly exhausted. Postwar changes in dengue epidemiology in the American region occurred somewhat later than in Asia. During the 1960s, dengue virus types 2 and 3 became established in the region, and in 1977, dengue type 1 was introduced, rapidly spread, and became endemic. The pattern of intermittent epidemics at long intervals and transient circulation of one serotype changed to one of annual outbreaks in multiple locations and persistent cocirculation of multiple dengue serotypes. It was predictable that these events would eventually lead to the emergence of DHF in the Americas (2). In the 1970s, a few DHF cases were identified in Puerto Rico, where dengue was under intense study. The first true outbreak occurred in Cuba in 1981, with 116,000 hospitalized patients, 34,000 documented cases of DHF, and 158 deaths (15). Similar events occurred in Venezuela in 1989–1990 (16), with over 3000 cases of hemorrhagic fever, and in Rio de Janeiro in 1990. During the decade that followed the Cuban epidemic, 11 countries in the Americas have reported DHF (Figure 3).
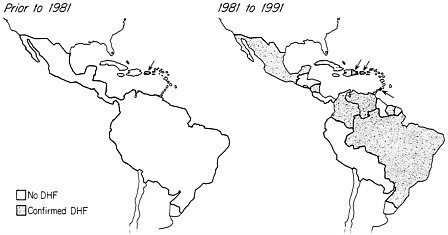
FIGURE 3 Expansion of DHF in the Western Hemisphere. Prior to 1981, only Puerto Rico reported sporadic cases of DHF. The first major outbreak occurred in Cuba in 1981. By 1991, 11 countries had been affected by the disease. Arrows indicate island nations affected in the Caribbean region.
DENGUE IN THE DEVELOPED WORLD
Every year 30–100 cases of dengue are reported in the United States in persons who have traveled to tropical countries. Many such cases undoubtedly go unreported. A similar incidence has been recorded among Swedish tourists, and it probably occurs elsewhere in Europe as well. Unlike most of Europe, parts of the United States have a resident fauna of efficient vectors (A. aegypti and Aedes albopictus; see below), and there is a risk of secondary spread. On two occasions (1980 and 1986) small outbreaks followed the introduction of dengue from Mexico into southern Texas. The introduction and spread of dengue outbreaks in the southern United States remains a potential threat, particularly in cities along the Gulf of Mexico, where A. aegypti and A. albopictus are abundant and major pest problems. As would be expected from the requirement for sequential infections, DHF has been rare in travelers and does not currently pose a threat to the developed world.
ECOLOGIC BASIS FOR THE EMERGENCE OF DENGUE
Underlying the emergence of DHF in the Western Hemisphere are changes in human and mosquito ecologies that affect the rate and geographic range of virus transmission. The principal vector, A. aegypti, has made extraordinary evolutionary adjustments to coexist with human
beings. This mosquito originated (and still exists) in Africa as a forest species feeding principally on rodents and other wild animals and adapted to lay eggs and undergo larval development in forest tree holes containing rainwater. However, a subspecies A. aegypti aegypti, evolved in Africa to become a highly domesticated animal, following humankind on its journeys and migrations to the corners of the globe, breeding in the artificial containers used for storing clean water, resting between blood meals in human habitations away from predators and harsh weather, flying rarely more than 50 yards from these convenient locations, and adopting wary biting habits around its observant and dexterous human prey. Interestingly, A. aegypti is not a very efficient vector of dengue viruses, and has both a low susceptibility to oral infection and low rate of transovarial infection (7). Thus, virus titers in the blood of human hosts must exceed 105–107 virus particles per ml for infection and transmission to be sustained. The vector may thus serve as an important selection mechanism or biological filter for maintaining virus virulence at a high level, since only virus strains that replicate efficiently in humans and produce high viremias are transmissible by this mosquito.
The phenomena of unchecked human population growth in the tropics and the dramatic redistribution of the human population into urban centers in search of better amenities, employment, and education have greatly influenced the epidemiology of dengue and the density and distribution of the domestic vector, A. aegypti. This redistribution of the human population has occurred in all countries where dengue is endemic. In the American region, for example, the urban population nearly doubled during the period (1970–1990) in which dengue emerged as a major health problem, whereas rural populations remained nearly constant. This trend will continue, with over 80% of the Latin American population predicted to inhabit cities within the next 30 years. Insufficient urban piped water supplies, necessitating the storage of water for drinking and washing, and poor sanitation, resulting in the accumulation of vast amounts of human detritus that collect rainwater, such as discarded bottles, cans, and automobile tires, have been responsible for an enormous expansion of A. aegypti vector populations. This environmental transformation has occurred in a setting where the supply of susceptible human hosts for dengue transmission is now virtually inexhaustible.
These changes in human and vector ecology have been accompanied by a decreased willingness of societies to undertake effective mosquito control in the context of disease prevention. In 1947, the Pan American Health Organization initiated a hemisphere-wide campaign to eradicate A. aegypti. Efforts were successful in a number of countries, with the
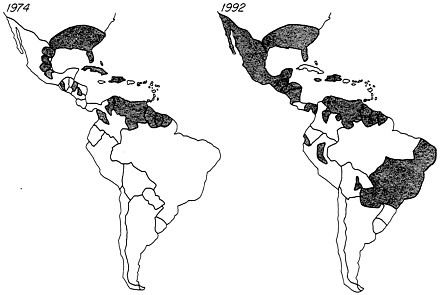
FIGURE 4 Reinfestation of Central and South America by A. aegypti (shaded areas) after several decades, ending in 1972, of intensive efforts to control this vector species. The reinfestation has led to explosive epidemics of dengue and DHF in South America.
result that between 1947 and 1972, the vector had been eliminated from 19 countries, representing over 73% of the area originally infested (Figure 4) (2, 17). However, around 1972 the program ran out of steam. The United States entered the campaign late (1961) and gave up early, as entomologists and sanitarians found the job insurmountable and congressmen found the funding requirements unacceptable. The United States and other countries in the Caribbean region provided a source for exportation of A. aegypti to countries attempting to maintain their vector-free status. In those countries that had achieved eradication, mosquito control efforts sagged under the competition with other priorities for scarce health resources and with the increasing demands imposed by the conditions in bursting megacities such as Sao Paulo and Rio de Janeiro. Lack of funding, leadership, and morale eroded the once-proud mosquito control agencies (18). The expansion of urban human populations in Latin America and the creation of vast amounts of nonbiodegradable detritus breeding vast populations of A. aegypti simply outstripped the human capability to restrict this species. Within 10 years A. aegypti had retaken virtually all of South and Central America (2) (Figure 4). In 1986, an explosive outbreak of dengue 1 involving over 1 million cases struck Rio de Janeiro. Subsequently, dengue epidemics
have occurred in Paraguay, Bolivia, Peru, Ecuador, Colombia, and Venezuela. The sequential introduction of dengue type 1 followed by dengue 2 resulted in outbreaks of DHF in Caracas (1989) and Rio de Janeiro (1990). Three of the four dengue serotypes are currently endemic in the region, and it is probable that dengue type 3 virus will be reintroduced in the near future. It is clear that the factors that led to the emergence of DHF as a major public health problem in Asia now exist in the American region.
INTRODUCTION AND SPREAD OF A SECOND DENGUE VECTOR IN THE AMERICAS
An interesting illustration of the relationship between human and vector ecology is provided by the introduction and spread of another dengue vector, A. albopictus, from Asia to the Americas in the 1980s. This mosquito is responsible for endemic transmission of dengue in Asia and for epidemic spread in circumstances where A. aegypti is absent or in low density. In 1985, the mosquito was discovered in Houston, Texas, and a year later in Rio de Janeiro, representing separate introductions from different areas of Asia (19, 20). Human commerce in used truck tires imported for the purpose of recapping was responsible for the importation of eggs or larvae of the mosquito (21). These tires were stored in the open prior to export, where they collected rainwater and were used as oviposition sites by A. albopictus. Over a million tires per year were imported into the United States from Asia, and approximately 20% of these, unfit for recapping purposes, were discarded in the environment. The A. albopictus invasion rapidly spread, probably in large part by the tire trade, extending the range of this winter-hardy species throughout the eastern United States. A similar expansion has occurred in Brazil. Dissemination of the mosquito is continuing, and in 1993 it invaded the Dominican Republic (C. Peña, personal communication). It was predicted that this aggressive and adaptable species would become implicated in the transmission of indigenous viruses, and this has in fact occurred; five different agents, including two human pathogens (eastern equine encephalitis and dengue viruses) have now been recovered from the species (ref. 22; P. Reiter, personal communication). The full dynamics of this unique episode have yet to play out, but it will be surprising if a significant public health problem does not emerge in the future. The accumulation of vast numbers of nonbiodegradable transportable man-made mosquito-breeding devices lies at the root of the dengue vector problem. Municipal landfills reject these objects, which are thus often dumped illegally in the environment, and no recycling has proven cost effective.
MOLECULAR EPIDEMIOLOGY AND VIRUS VARIATION
Little is presently known about the role of virus-specified factors in the transmission and pathogenesis of dengue. Considerable microevolution of dengue virus strains has been found in studies employing monoclonal antibody analysis (23), RNA fingerprinting (24), or sequencing of selected regions of the genome (25). Since variants with similar genetic structure are found within a specific geographic location, such studies have elucidated movements of dengue viruses and the source of epidemics. The results also suggest that genetic variation in virus strains may determine virulence and explain the changing patterns of disease. Two examples will be given to illustrate these different points.
Use of Dengue Genetic Analysis to Explain the Emergence of Dengue in Africa
In modern times, there were virtually no reports of human dengue in Africa except for scattered cases in the 1960s in Nigeria and 1970s in Senegal. Serological surveys in 1970 in Nigeria suggested the presence of a zoonotic transmission cycle involving monkeys and a variety of tree-hole-breeding Aedes spp. (26), and between 1974 and 1981, French scientists verified the existence of such a cycle in Côte d'Ivoire and Senegal (27). However, no outbreaks of human disease were recorded in the region around forests supporting this cycle, and the relevance of a forest transmission cycle to human health remains obscure. Indeed, the outbreak of dengue fever closest to Africa occurred in the Indian Ocean (Seychelle Islands) in 1977.
In 1982, however, dengue appeared along the coast of Kenya, and shortly thereafter in the Ivory Coast and Burkina Faso. These were typical A. aegypti-borne epidemics, characterized by classical dengue fever. In 1983–1984, the disease spread up the coast of East Africa to involve Sudan and Somalia, where it has remained endemic. These events posed a number of questions: Why did these outbreaks arise? Were they interconnected? Was there a relationship between the outbreaks and the indigenous forest transmission cycle? These questions were resolved by a comparison of strains using hybridization probes (28) and by an analysis of nucleotide sequences of a 240-bp region at the junction of the E and NS1 proteins (25). Virus strains were available for study representing the forest cycle and the epidemics in Burkina Faso and the Seychelles. A close genetic relationship was found between the strains from West Africa representing the forest cycle, and these strains were quite distinct from those causing the epidemics. The virus strains from the Burkina Faso epidemic were similar to the strains responsible
for the earlier outbreak in the Seychelles, and these in turn resembled virus isolates from Sri Lanka and Indonesia. Thus, it appeared that the epidemics arose by introduction of dengue 2 virus from afar and were not the result of spill-over from the forest transmission cycle. The molecular analysis confirmed the role of humans in the dissemination of dengue viruses. The virus was apparently introduced to the Seychelles in 1977 from Sri Lanka or another location to the East, and this was also the source of introduction into East and West Africa.
Molecular and Biological Evidence for Variation in Dengue Virulence
In the Americas, dengue type 2 has been responsible for repeated outbreaks of classical dengue fever as well as for the explosive appearance of DHF in Cuba, Venezuela, and Brazil. This paradox could be explained solely on the basis of the epidemiological events leading to the establishment of hyperendemic transmission of multiple serotypes and an increased incidence of sequential infections. However, circumstantial evidence from molecular analysis suggests that the dengue 2 strain responsible for the occurrence of severe disease may represent a variant with increased virulence for humans.
A comparison of the gene sequences of dengue type 2 viruses from the Caribbean region indicated that two distinct variants were cocirculating over a period of many years (25). One genetic variant represented the Puerto Rican strain, which had been introduced into the region in 1969 and had persisted in an endemic-epidemic pattern, associated with classical dengue fever. A second variant represented a strain first isolated in Jamaica in 1981. The Jamaican genotype was responsible for the DHF epidemics in Venezuela (1989) and Brazil (1990). Since Jamaica and Cuba are geographically juxtaposed, it is probable that this variant caused the 1981 Cuban DHF outbreak (virus strains isolated in Cuba have never been available for comparative study).* Sequence homology between the Jamaican variant and virus strains from Vietnam suggested the original source of introduction—a plausible conclusion, since Cuba
and North Vietnam had political ties at the time of the outbreak. In contrast, the Puerto Rican genotype resembled contemporary virus strains from Polynesia, where for the most part disease was mild.
Although these results imply that dengue virus strains vary in virulence and that this variation may play a role in the incidence and distribution of DHF, the molecular basis for virulence cannot be tested directly, due to the lack of an animal model of DHF. A preliminary analysis (29) demonstrating variation in the ability of dengue virus strains to replicate in human peripheral blood monocytes and higher replication of isolates from DHF patients than from dengue fever patients requires further evaluation and confirmation in prospective studies.
FLAVIVIRUS AND DENGUE EVOLUTION:A BIOLOGICAL PERSPECTIVE AND SOME SPECULATIONSON FUTURE CHANGES IN DENGUE EPIDEMIOLOGY
The 68 recognized flaviviruses are classified into serological complexes, and these in turn correspond closely to their mode of transmission. Nearly half, including the dengue viruses, are mosquito-borne, 28% are transmitted by ticks, and 20% are apparently zoonotic infections transmitted by contact between rodents and bats (30). From an evolutionary standpoint, it is of interest that the viruses belonging to the tick-borne encephalitis complex seem to have developed the capacity to infect epithelial surfaces, since members of this group are shed in milk of infected domestic livestock and can spread to humans by this means as well as by aerosols in the laboratory. This capacity for epithelial infection is apparently more highly developed in the bat- and rodent-associated agents which have no known arthropod vectors, probably representing an adaptation to transmission between hosts that are relatively abundant under circumstances of low vector density. Evolution of the flaviviruses appears to be in the direction of loss of dependence on arthropod vectors, with the most evolved forms represented by hepatitis C and simian hemorrhagic fever viruses.
The mosquito-borne flaviviruses, exemplified by St. Louis encephalitis, Japanese encephalitis, and Murray Valley encephalitis viruses, are presumed to be relatively primitive forms on the basis of their varied vector and host associations and marked radiation of closely related antigenic types. A biological common denominator of these agents is their marked neurotropism. Dengue and yellow fever viruses are distinct antigenically but share links with the more primitive mosquito-borne encephalitis viruses, as illustrated by the high nucleotide sequence homology between dengue type 2 virus and Edge Hill virus (31).
However, dengue and yellow fever viruses have evolved an important biological feature that clearly separates them from their more primitive neurotropic predecessors, namely lymphotropism, and they have subverted the primate monocyte/macrophage as principal target cell for replication (32). One result of this adaptation has been a marked restriction of host range to the primates and a parallel restriction in the virus-vector relationship to a few species of Aedes spp. with predilection for feeding on primate blood.
The lymphotropism of dengue viruses is consistent with the production and release of large quantities of virus into the blood stream, a feature that is essential to transmission of the virus by the relatively insusceptible A. aegypti vector. It is important to note that the change in tropism from nervous tissue to lymphoid cells is critical to survival of both virus and host species, since viremias of the magnitude achieved in dengue would produce a very high risk of hematogenous spread of a neurovirulent/neuroinvasive agent to the central nervous system. Loss of neurovirulence in favor of lymphotropism is thus permissive for transmission without affecting host survival.
The remarkable ability of dengue viruses to replicate to high titers in human tissues raises the possibility that, should a variant emerge with altered tropism—e.g., for epithelial tissues permitting shedding in respiratory secretions, or conversely for nervous tissues, permitting neuroinvasion—the result could fundamentally change the route of transmission or the expression of disease in the host. While this may appear to be a remote possibility, we should remember that other flaviviruses, including the bat- and rodent-associated flaviviruses and simian hemorrhagic fever virus, have evolved in these directions. It is thus not inconceivable in the context of flavivirus evolution that dengue could undergo genetic changes that would transform it into a more dangerous pathogen transmissible by the respiratory route or capable of causing encephalitis.
In general, however, dengue and other flaviviruses [e.g., yellow fever virus (33) and St. Louis encephalitis virus (34)] display a rather remarkable genetic stability, given their RNA genomes and high rates of mutation. A comparison of the genetic variability of dengue type 2 strains with that of influenza viruses, based on a comparison of nucleotide and amino acid sequence divergence, shows that dengue variation is low, similar to that of influenza C, a virus that has a single (human) host and is not subjected to the marked genetic shifts typical of influenza A virus, which undergoes recombinational events in alternative hosts and has a higher proportion of mutations that result in amino acid changes. Dengue virus strains belonging to the same serotype vary by no more than 10% at the nucleotide level and 4% at the amino acid
PREVENTION AND CONTROL
The ability to control dengue by reduction of the vectors responsible for transmission seems increasingly remote. Efforts to develop effective live, attenuated, and genetically engineered vaccines are under way, and the impetus to move candidate vaccines into large-scale human trials will increase as DHF emerges as a major health problem in the Western Hemisphere. The immunopathogenesis of DHF demands that durable protective immunity to all four serotypes be elicited simultaneously, posing a formidable challenge to vaccine development.
SUMMARY
Dengue viruses are members of the Flaviviridae, transmitted principally in a cycle involving humans and mosquito vectors. In the last 20 years the incidence of dengue fever epidemics has increased and hyperendemic transmission has been established over a geographically expanding area. A severe form, dengue hemorrhagic fever (DHF), is an immunopathologic disease occurring in persons who experience sequential dengue infections. The risk of sequential infections, and consequently the incidence of DHF, has risen dramatically, first in Asia and now in the Americas. At the root of the emergence of dengue as a major health problem are changes in human demography and behavior, leading to unchecked populations of and increased exposure to the principal domestic mosquito vector, Aedes aegypti. Virus-specified factors also influence the epidemiology of dengue. Speculations on future events in the epidemiology, evolution, and biological expression of dengue were presented.
References
1. Halstead, S. B. (1980) Bull. W.H.O. 58, 1–21.
2. Gubler, D. J. (1989) Am. J. Trop. Med. Hyg. 40, 571–578.
3. Pinheiro, F. P. (1989) Epidemiol. Bull. (PAHO) 10, 1–8.
4. Gubler, D. J. (1991) Virus Info. Exchange Newslett. 8, 2–3.
5. Chambers, T. J., Hahn, C. S. & Galler, R. (1990) Annu. Rev. Microbiol . 44, 649.
6. Rudnick, A. (1978) Asian J. Infect. Dis. 2, 156–168.
7. Gubler, D. J. (1988) in The Arboviruses: Epidemiology and Ecology , ed. Monath, T. P. (CRC, Boca Raton, FL), Vol. 2, pp. 223–260.
8. Halstead, S. B. (1989) Rev. Infect. Dis. 11, S830–S839.
9. Halstead, S. B. (1988) Science 239, 476–481.
10. Kurane, I. & Ennis, F. A. (1992) Semin. Immunol. 4, 121–130.
11. Carey, D. E. (1971) J. Hist. Med. 26, 243–262.
12. Ehrenkrantz, N. J., Ventura, A. K., Cuadrado, R. R., Pond, W. L. & Porter, J. E. (1971) N. Engl. J. Med. 285, 1460–1469.
13. Gubler, D. J. (1989) in Dengue Hemorrhagic Fever: Diagnosis, Treatment and Control, ed. Thongcharoen, P. (W.H.O., New Delhi).
14. Halstead, S. B. (1990) Asian J. Trop. Med. Pub. Health 21, 636.
15. Kouri, G. & Guzman, M. G. (1986) Bull. PAHO 20, 24–26.
16. Anonymous (1990) Epidemiol. Bull. (PAHO) 2, 1–2.
17. Schliesman, D. J. & Calheiros, L. B. (1974) Mosq. News 34, 1–9.
18. Halstead, S. B. (1984) Rev. Infect. Dis. 16, 251–264.
19. Sprenger, D. & Wuithiranyagool, T. (1986) Am. Mosq. Control Assoc. J. 2, 217–221.
20. Forattini, O. P. (1986) Rev. Saude Publica 20, 244–250.
21. Reiter, P. & Sprenger, D. (1987) Am. Mosq. Control Assoc. J. 3, 494–500.
22. Mitchell, C. J., Niebylski, M. L., Smith, G. C., Karabatsos, N., Martin, D., Mutebi, J.-P., Craig, G. B., Jr., & Mahler, M. J. (1992) Science 257, 526–527.
23. Monath, T. P., Wands, J. R., Hill, L. J., Brown, N. V., Marciniak, R. A., Wong, M. A., Gentry, M. K., Burke, D. S., Grant, J. A. & Trent, D. W. (1986) Virology 154, 313–324.
24. Trent, D. W., Grant, J. A., Rosen, L. & Monath, T. P. (1983) Virology 128, 271–284.
25. Rico-Hesse, R. (1990) Virology 174, 479–493.
26. Monath, T. P., Lee, V. H., Wilson, D. C., Fagbami, A. & Tomori, O. (1974) Trans. R. Soc. Trop. Med. Hyg. 68, 30–39.
27. Roche, J. C., Cordellier, R., Hervy, J. P., Digoutte, J. P. & Monteny, N. (1983) Ann. Virol. 134, 233–239.
28. Kerschner, J. A., Vorndam, A. V., Monath, T. P. & Trent, D. W. (1986) J. Gen. Virol. 67, 2645–2661.
29. Kliks, S. (1992) AIDS Res. Hum. Retroviruses 6, 993–998.
30. Karabatsos, N., ed. (1985) Catalogue of Arthropod-Borne Viruses and Certain Other Viruses of Vertebrates (Am. Soc. Trop. Med. Hyg., San Antonio, TX).
31. Blok, J., Henchal, E. A. & Gorman, B. J. (1984) J. Gen. Virol. 65, 2173–2180.
32. Monath, T. P. (1985) in The Togaviridae and Flaviviridae, eds. Schlesinger, S. & Schlesinger, M. J. (Plenum, New York), pp. 375–440.
33. Duebel, V., Digoutte, J. P., Monath, T. P. & Girard, M. (1986) J. Gen. Virol. 67, 209–213.
34. Trent, D. W., Monath, T. P, Bowen, G. S., Vorndam, A. V., Cropp, C. B. & Kemp, G. E. (1980) Ann. N.Y. Acad. Sci. 354, 219–228.
35. Gubler, D. J. (1985) Am. J. Trop. Med. Hyg. 34, 170–173.
















