
Evidence Supporting Biological Evolution
Along path leads from the origins of primitive "life," which existed at least 3.5 billion years ago, to the profusion and diversity of life that exists today. This path is best understood as a product of evolution.
Contrary to popular opinion, neither the term nor the idea of biological evolution began with Charles Darwin and his foremost work, On the Origin of Species by Means of Natural Selection (1859). Many scholars from the ancient Greek philosophers on had inferred that similar species were descended from a common ancestor. The word "evolution" first appeared in the English language in 1647 in a nonbiological connection, and it became widely used in English for all sorts of progressions from simpler beginnings. The term Darwin most often used to refer to biological evolution was "descent with modification," which remains a good brief definition of the process today.
Darwin proposed that evolution could be explained by the differential survival of organisms following their naturally occurring variation—a process he termed "natural selection." According to this view, the offspring of organisms differ from one another and from their parents in ways that are heritable—that is, they can pass on the differences genetically to their own offspring. Furthermore, organisms in nature typically produce more offspring than can survive and reproduce given the constraints of food, space, and other environmental resources. If a particular off-

Charles Darwin arrived at many of his insights into evolution by studying the variations among species on the Galápagos Islands off the coast of Ecuador.

spring has traits that give it an advantage in a particular environment, that organism will be more likely to survive and pass on those traits. As differences accumulate over generations, populations of organisms diverge from their ancestors.
Darwin's original hypothesis has undergone extensive modification and expansion, but the central concepts stand firm. Studies in genetics and molecular biology—fields unknown in Darwin's time—have explained the occurrence of the hereditary variations that are essential to natural selection. Genetic variations result from changes, or mutations, in the nucleotide sequence of DNA, the molecule that genes are made from. Such changes in DNA now can be detected and described with great precision.
Genetic mutations arise by chance. They may or may not equip the organism with better means for surviving in its environment. But if a gene variant improves adaptation to the environment (for example, by allowing an organism to make better use of an available nutrient, or to escape predators more effectively—such as through stronger legs or disguising coloration), the organisms carrying that gene are more likely to survive and reproduce than those without it. Over time, their descendants will tend to increase, changing the average characteristics of the population. Although the genetic variation on which natural selection works is based on random or chance elements, natural selection itself produces "adaptive" change—the very opposite of chance.
Scientists also have gained an understanding of the processes by which new species originate. A new species is one in which the individuals cannot mate and produce viable descendants with individuals of a preexisting species. The split of one species into two often starts because a group of individuals becomes geographically separated from the rest. This is particularly apparent in distant remote islands, such as the Galápagos and the Hawaiian archipelago, whose great distance from the Americas and Asia means that arriving colonizers will have little or no opportunity to mate with individuals remaining on those continents. Mountains, rivers, lakes, and other natural barriers also account for geographic separation between populations that once belonged to the same species.
Once isolated, geographically separated groups of individuals become genetically differentiated as a consequence of mutation and other processes, including natural selection. The origin of a species is often a gradual process, so that at first the reproductive isolation between separated groups of organisms is only partial, but it eventually becomes complete. Scientists pay special attention to these intermediate situations, because they help to reconstruct the details of the process and to identify particular genes or sets of genes that account for the reproductive isolation between species.
A particularly compelling example of speciation involves the 13 species of finches studied by Darwin on the Galápagos Islands, now known as Darwin's finches. The ancestors of these finches appear to have immigrated from the South American mainland to the Galápagos. Today the different species of finches on the island have distinct habitats, diets, and behaviors, but the mechanisms involved in speciation continue to operate. A research group led by Peter and Rosemary Grant of Princeton University has shown that a single year of drought on the islands can drive evolutionary changes in the finches. Drought diminishes supplies of easily

The different species of finches on the Galápagos Islands, now known as Darwin's finches, have different-sized beaks that have evolved to take advantage of distinct food sources.
cracked nuts but permits the survival of plants that produce larger, tougher nuts. Droughts thus favor birds with strong, wide beaks that can break these tougher seeds, producing populations of birds with these traits. The Grants have estimated that if droughts occur about once every 10 years on the islands, a new species of finch might arise in only about 200 years.
The following sections consider several aspects of biological evolution in greater detail, looking at paleontology, comparative anatomy, biogeography, embryology, and molecular biology for further evidence supporting evolution.
The Fossil Record
Although it was Darwin, above all others, who first marshaled convincing evidence for biological evolution, earlier scholars had recognized that organisms on Earth had changed systematically over long periods of time. For example, in 1799 an engineer named William Smith reported that, in undisrupted layers of rock, fossils occurred in a definite sequential order, with more modern-appearing ones closer to the top. Because bottom layers of rock logically were laid down earlier and thus are older than top layers, the sequence of fossils also could be given a chronology from oldest to youngest. His findings were confirmed and extended in the 1830s by the paleontologist William Lonsdale, who recognized that fossil remains of organisms from lower strata were more primitive than the ones above. Today, many thousands of ancient rock deposits have been identified that show corresponding successions of fossil organisms.
Thus, the general sequence of fossils had already been recognized before Darwin conceived of descent with modification. But the paleontologists and geologists before Darwin used the sequence of fossils in rocks not as proof of biological evolution, but as a basis for working out the original sequence of rock strata that had been structurally disturbed by earthquakes and other forces.
In Darwin's time, paleontology was still a rudimentary science. Large parts of the geological succession of stratified rocks were unknown or inadequately studied.
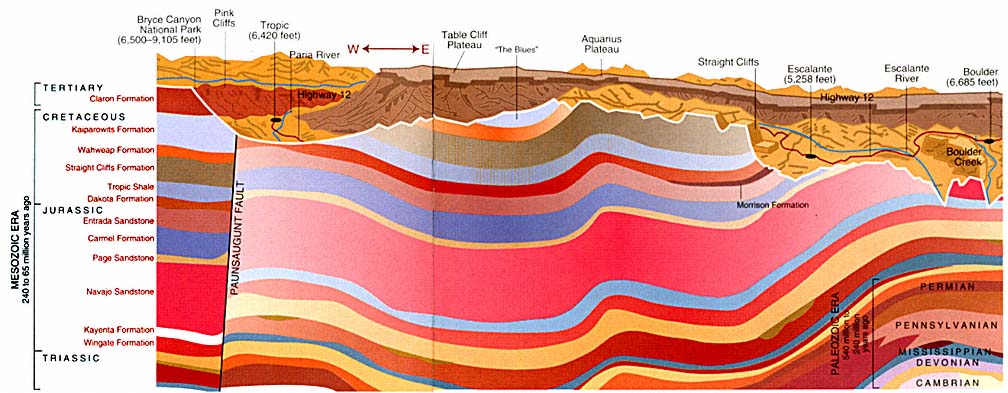
A geological cross section of the Grand Staircase-Escalante National Monument in Utah shows layers of sedimentary rock. These layers reveal deposits laid down over millions of years. Older fossils are found in the lower layers, revealing the succession of organisms over time

Weathering has exposed layers of sedimentary rock near the Paria River in Utah
Darwin, therefore, worried about the rarity of intermediate forms between some major groups of organisms.
Today, many of the gaps in the paleontological record have been filled by the research of paleontologists. Hundreds of thousands of fossil organisms, found in well-dated rock sequences, represent successions of forms through time and manifest many evolutionary transitions. As mentioned earlier, microbial life of the simplest type was already in existence 3.5 billion years ago. The oldest evidence of more complex organisms (that is, eucaryotic cells, which are more complex than bacteria) has been discovered in fossils sealed in rocks approximately 2 billion years old. Multicellular organisms, which are the familiar fungi, plants, and animals, have been found only in younger geological strata. The following list presents the order in which increasingly complex forms of life appeared:
|
Life Form |
Millions of Years Since First Known Appearance (Approximate) |
|
Microbial (procaryotic cells) |
3,500 |
|
Complex (eucaryotic cells) |
2,000 |
|
First multicellular animals |
670 |
|
Shell-bearing animals |
540 |
|
Vertebrates (simple fishes) |
490 |
|
Amphibians |
350 |
|
Reptiles |
310 |
|
Mammals |
200 |
|
Nonhuman primates |
60 |
|
Earliest apes |
25 |
|
Australopithecine ancestors of humans |
5 |
|
Modern humans |
0.15 (150,000 years) |
So many intermediate forms have been discovered between fish and amphibians, between amphibians and reptiles, between reptiles and mammals, and along the primate lines of descent that it often is difficult to identify categorically when the transition occurs from one to another particular species. Actually, nearly all fossils can be regarded as intermediates in some sense; they are life forms that come between the forms that preceded them and those that followed.
The fossil record thus provides consistent evidence of systematic change through time—of descent with modification. From this huge body of evidence, it can be predicted that no reversals will be found in future paleontological studies. That is, amphibians will not appear before fishes, nor mammals before reptiles, and no complex life will occur in the geological record before the oldest eucaryotic cells. This prediction has been upheld by the evidence that has accumulated until now: no reversals have been found.
Common Structures
Inferences about common descent derived from paleontology are reinforced by comparative anatomy. For example, the skeletons of humans, mice, and bats are strikingly similar, despite the different ways of life of these animals and the diversity of environments in which they flourish. The correspondence of these animals, bone by bone, can be observed in every part of the body, including the limbs; yet a person writes, a mouse runs, and a bat flies with structures built of bones that are different in detail but similar in general structure and relation to each other.
Scientists call such structures homologies and have concluded that they are best explained by common descent. Comparative anatomists investigate such homologies, not only in bone structure but also in other parts of the body, working out relationships from degrees of similarity. Their conclusions provide important inferences about the details of evolutionary history, inferences that can be tested by comparisons with the sequence of ancestral forms in the paleontological record.
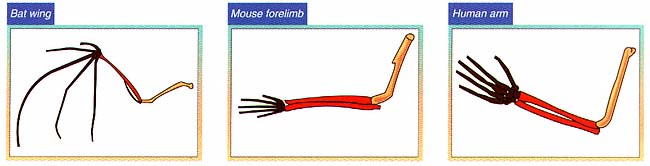
A bat wing, a mouse forelimb, and a human arm serve very different purposes, but they have the same basic components The similarities arise because all three species share a common four-limbed vertebrate ancestor
The mammalian ear and jaw are instances in which paleontology and comparative anatomy combine to show common ancestry through transitional stages. The lower jaws of mammals contain only one bone, whereas those of reptiles have several. The other bones in the reptile jaw are homologous with bones now found in the mammalian ear. Paleontologists have discovered intermediate forms of mammal-like reptiles (Therapsida) with a double jaw joint—one composed of the bones that persist in mammalian jaws, the other consisting of bones that eventually became the hammer and anvil of the mammalian ear.
The Distribution of Species
Biogeography also has contributed evidence for descent from common ancestors. The diversity of life is stupendous. Approximately 250,000 species of living plants, 100,000 species of fungi, and one million species of animals have been described and named, each occupying its own peculiar ecological setting or niche; and the census is far from complete. Some species, such as human beings and our companion the dog, can live under a wide range of environments. Others are amazingly specialized. One species of a fungus (Laboulbenia) grows exclusively on the rear portion of the covering wings of a single species of beetle (Aphaenops cronei) found only in some caves of southern France. The larvae of the fly Drosophila carcinophila can develop only in specialized grooves beneath the flaps of the third pair of oral appendages of a land crab that is found only on certain Caribbean islands.
How can we make intelligible the colossal diversity of living beings and the existence of such extraordinary, seemingly whimsical creatures as the fungus, beetle, and fly described above? And why are island groups like the Galápagos so often inhabited by forms similar to those on the nearest mainland but belonging to different species? Evolutionary theory explains that biological diversity results from the descendants of local or migrant predecessors becoming adapted to their diverse environments. This explanation can be tested by examining present species and local fossils to see whether they have similar structures, which would indicate how one is derived from the other. Also, there should be evidence that species without an established local ancestry had migrated into the locality.
Wherever such tests have been carried out, these conditions have been confirmed. A good example is provided by the mammalian populations of North and South America, where strikingly different native organisms evolved in isolation until the emergence of the isthmus of Panama approximately 3 million years ago. Thereafter, the armadillo, porcupine, and opossum—mammals of South American origin—migrated north, along with many other species of plants and animals, while the mountain lion and other North American species made their way across the isthmus to the south.
The evidence that Darwin found for the influence of geographical distribution on the evolution of organisms has become stronger with advancing knowledge. For example, approximately 2,000 species of flies belonging to the genus Drosophila are now found throughout the world. About one-quarter of them live only in Hawaii.


Until about 3 million years ago, North and South America were separated by a wide expanse of water, so mammals on the two continents evolved separately. After the isthmus of Panama formed, armadillos and opossums migrated north, and mountain lions migrated south. These movements are documented in the fossil record.
More than a thousand species of snails and other land mollusks also are found only in Hawaii. The biological explanation for the multiplicity of related species in remote localities is that such great diversity is a consequence of their evolution from a few common ancestors that colonized an isolated environment. The Hawaiian Islands are far from any mainland or other islands, and on the basis of geological evidence they never have been attached to other lands. Thus, the few colonizers that reached the Hawaiian Islands found many available ecological niches, where they could, over numerous generations, undergo evolutionary change and diversification. No mammals other than one bat species lived in the Hawaiian Islands when the first human settlers arrived; similarly, many other kinds of plants and animals were absent.
The Hawaiian Islands are not less hospitable than other parts of the world for the absent species. For example, pigs and goats have multiplied in the wild in Hawaii, and other domestic animals also thrive there. The scientific explanation for the absence of many kinds of organisms, and the great multiplication of a few kinds, is that many sorts of organisms never reached the islands, because of their geographic isolation. Those that did reach the islands diversified over time because of the absence of related organisms that would compete for resources.
Similarities During Development
Embryology, the study of biological development from the time of conception, is another source of independent evidence for common descent. Barnacles, for instance, are sedentary crustaceans with little apparent similarity to such other crustaceans as lobsters, shrimps, or copepods. Yet barnacles pass through a free-swimming larval stage in which they look like other crustacean larvae. The similarity of larval stages supports the conclusion that all crustaceans have homologous parts and a common ancestry.
Similarly, a wide variety of organisms from fruit flies to worms to mice to humans have very similar sequences of genes that are active early in development. These genes influence body segmentation or orientation in all these diverse groups. The presence of such similar genes doing similar things across such a wide range of organisms is best explained by their having been present in a very early common ancestor of all of these groups.
New Evidence from Molecular Biology
The unifying principle of common descent that emerges from all the foregoing lines of evidence is being reinforced by the discoveries of modern biochemistry and molecular biology.
The code used to translate nucleotide sequences into amino acid sequences is essentially the same in all organisms. Moreover, proteins in all organisms are invariably composed of the same set of 20 amino acids. This unity of composition
and function is a powerful argument in favor of the common descent of the most diverse organisms.
In 1959, scientists at Cambridge University in the United Kingdom determined the three-dimensional structures of two proteins that are found in almost every multicelled animal: hemoglobin and myoglobin. Hemoglobin is the protein that carries oxygen in the blood. Myoglobin receives oxygen from hemoglobin and stores it in the tissues until needed. These were the first three-dimensional protein structures to be solved, and they yielded some key insights. Myoglobin has a single chain of 153 amino acids wrapped around a group of iron and other atoms (called "heme") to which oxygen binds. Hemoglobin, in contrast, is made of up four chains: two identical chains consisting of 141 amino acids, and two other identical chains consisting of 146 amino acids. However, each chain has a heme exactly like that of myoglobin, and each of the four chains in the hemoglobin molecule is folded exactly like myoglobin. It was immediately obvious in 1959 that the two molecules are very closely related.
During the next two decades, myoglobin and hemoglobin sequences were determined for dozens of mammals, birds, reptiles, amphibians, fish, worms, and molluscs. All of these sequences were so obviously related that they could be compared with confidence with the three-dimensional structures of two selected standards—whale myoglobin and horse hemoglobin. Even more significantly, the differences between sequences from different organisms could be used to construct a family tree of hemoglobin and myoglobin variation among organisms. This tree agreed completely with observations derived from paleontology and anatomy about the common descent of the corresponding organisms.
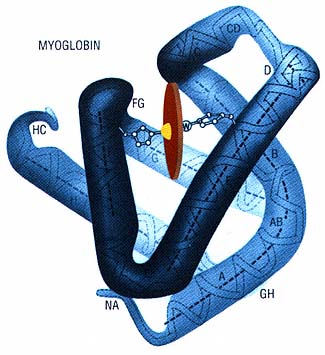
Myoglobin, which stores oxygen in muscles, consists of a chain of 153 amino acids wrapped around an oxygen-binding molecule. The sequence of amino acids in myoglobin vanes from species to species, revealing the evolutionary relationships among organisms.
Similar family histories have been obtained from the three-dimensional structures and amino acid sequences of other proteins, such as cytochrome c (a protein engaged in energy transfer) and the digestive proteins trypsin and chymotrypsin. The examination of molecular structure offers a new and extremely powerful tool for studying evolutionary relationships. The quantity of information is potentially huge—as large as the thousands of different proteins contained in living organisms, and limited only by the time and resources of molecular biologists.
As the ability to sequence the nucleotides making up DNA has improved, it also has become possible to use genes to reconstruct the evolutionary history of organisms. Because of mutations, the sequence of nucleotides in a gene gradually changes over time. The more closely related two organisms are, the less different their DNA will be. Because there are tens of thousands of genes in humans and other organisms, DNA contains a tremendous amount of information about the evolutionary history of each organism.
Genes evolve at different rates because, although mutation is a random event, some proteins are much more tolerant of changes in their amino acid sequence than
are other proteins. For this reason, the genes that encode these more tolerant, less constrained proteins evolve faster The average rate at which a particular kind of gene or protein evolves gives rise to the concept of a "molecular clock." Molecular clocks run rapidly for less constrained proteins and slowly for more constrained proteins, though they all time the same evolutionary events.
The figure on this page compares three molecular clocks: for cytochrome c proteins, which interact intimately with other macromolecules and are quite constrained in their amino acid sequences; for the less rigidly constrained hemoglobins, which interact mainly with oxygen and other small molecules; and for fibrinopeptides, which are protein fragments that are cut from larger proteins (fibrinogens) when blood clots. The clock for fibrinopeptides runs rapidly; 1 percent of the amino acids change in a little longer than 1 million years. At the other extreme, the molecular clock runs slowly for cytochrome c; a 1 percent change in amino acid sequence requires 20 million years. The hemoglobin clock is intermediate.
The concept of a molecular clock is useful for two purposes. It determines evolutionary relationships among organisms, and it indicates the time in the past when species started to diverge from one another. Once the clock for a particular gene or protein has been calibrated by reference to some event whose time is known, the actual chronological time when all other events occurred can be determined by examining the protein or gene tree.
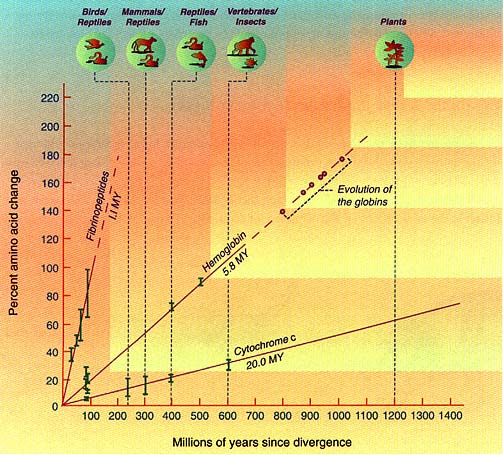
Species that diverged longer ago have more differences in their corresponding proteins, reflecting changes in the amino acids over time. Proteins evolve at different rates depending on the constraints imposed by their functions. Cytochrome c, a protein involved in energy transfer, is tightly constrained and changes slowly. Fibrinopeptides, which are involved in blood clotting, are much less constrained, with hemoglobin an intermediate case. The estimates for times of divergence shown here are based on 1971 data and have changed slightly since then (see table on page 13).
An interesting additional line of evidence supporting evolution involves sequences of DNA known as "pseudogenes." Pseudogenes are remnants of genes that no longer function but continue to be carried along in DNA as excess baggage. Pseudogenes also change through time, as they are passed on from ancestors to descendants, and they offer an especially useful way of reconstructing evolutionary relationships.
With functioning genes, one possible explanation for the relative similarity between genes from different organisms is that their ways of life are similar—for example, the genes from a horse and a zebra could be more similar because of their similar habitats and behaviors than the genes from a horse and a tiger. But this possible explanation does not work for pseudogenes, since they perform no function. Rather, the degree of similarity between pseudogenes must simply reflect their evolutionary relatedness. The more remote the last common ancestor of two organisms, the more dissimilar their pseudogenes will be.
The evidence for evolution from molecular biology is overwhelming and is growing quickly. In some cases, this molecular evidence makes it possible to go beyond the paleontological evidence. For example, it has long been postulated that whales descended from land mammals that had returned to the sea. From anatomical and paleontological evidence, the whales' closest living land relatives seemed to be the even-toed hoofed mammals (modem cattle, sheep, camels, goats, etc.).
Recent comparisons of some milk protein genes (beta-casein and kappa-casein) have confirmed this relationship and have suggested that the closest land-bound living relative of whales may be the hippopotamus. In this case, molecular biology has augmented the fossil record.
Creationism and the Evidence for Evolution
Some creationists cite what they say is an incomplete fossil record as evidence for the failure of evolutionary theory. The fossil record was incomplete in Darwin's time, but many of the important gaps that existed then have been filled by subsequent paleontological research. Perhaps the most persuasive fossil evidence for evolution is the consistency of the sequence of fossils from early to recent. Nowhere on
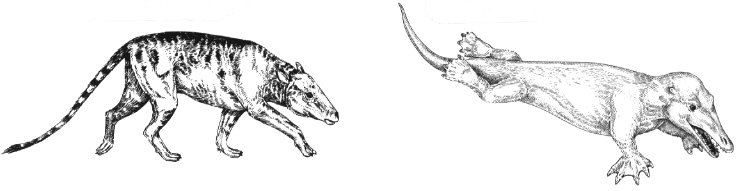
Mammakian land ancestor
Ambulocetus
Modern whales trace their ancestry to land mammals that evolved into species progressively more adapted to the water.
Earth do we find, for example, mammals in Devonian (the age of fishes) strata, or human fossils coexisting with dinosaur remains. Undisturbed strata with simple unicellular organisms predate those with multicellular organisms, and invertebrates precede vertebrates; nowhere has this sequence been found inverted. Fossils from adjacent strata are more similar than fossils from temporally distant strata. The most reasonable scientific conclusion that can be drawn from the fossil record is that descent with modification has taken place as stated in evolutionary theory.
Special creationists argue that "no one has seen evolution occur." This misses the point about how science tests hypotheses. We don't see Earth going around the sun or the atoms that make up matter. We "see" their consequences. Scientists infer that atoms exist and Earth revolves because they have tested predictions derived from these concepts by extensive observation and experimentation.
Furthermore, on a minor scale, we "experience" evolution occurring every day. The annual changes in influenza viruses and the emergence of antibiotic-resistant bacteria are both products of evolutionary forces. Indeed, the rapidity with which organisms with short generation times, such as bacteria and viruses, can evolve under the influence of their environments is of great medical significance. Many laboratory experiments have shown that, because of mutation and natural selection, such microorganisms can change in specific ways from those of immediately preceding generations.
On a larger scale, the evolution of mosquitoes resistant to insecticides is another example of the tenacity and adaptability of organisms under environmental stress. Similarly, malaria parasites have become resistant to the drugs that were used extensively to combat them for many years. As a consequence, malaria is on the increase, with more than 300 million clinical cases of malaria occurring every year.
Molecular evolutionary data counter a recent proposition called "intelligent design theory." Proponents of this idea argue that structural complexity is proof of the direct hand of God in specially creating organisms as they are today. These arguments echo those of the 18th century cleric William Paley who held that the vertebrate eye, because of its intricate organization, had been specially designed in its present form by an omnipotent Creator. Modem-day intelligent design proponents argue that molecular structures such as DNA, or molecular processes such as
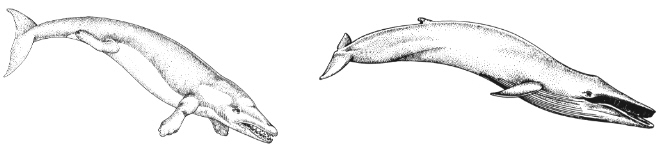
Rodhocetus
Balaenoptera (modern Blue whale)
the many steps that blood goes through when it clots, are so irreducibly complex that they can function only if all the components are operative at once. Thus, proponents of intelligent design say that these structures and processes could not have evolved in the stepwise mode characteristic of natural selection.
However, structures and processes that are claimed to be "irreducibly" complex typically are not on closer inspection. For example, it is incorrect to assume that a complex structure or biochemical process can function only if all its components are present and functioning as we see them today. Complex biochemical systems can be built up from simpler systems through natural selection. Thus, the "history" of a protein can be traced through simpler organisms. Jawless fish have a simpler hemoglobin than do jawed fish, which in turn have a simpler hemoglobin than mammals.
The evolution of complex molecular systems can occur in several ways. Natural selection can bring together parts of a system for one function at one time and then, at a later time, recombine those parts with other systems of components to produce a system that has a different function. Genes can be duplicated, altered, and then amplified through natural selection. The complex biochemical cascade resulting in blood clotting has been explained in this fashion.
Similarly, evolutionary mechanisms are capable of explaining the origin of highly complex anatomical structures. For example, eyes may have evolved independently many times during the history of life on Earth. The steps proceed from a simple eye spot made up of light-sensitive retinula cells (as is now found in the flatworm), to formation of individual photosensitive units (ommatidia) in insects with light focusing lenses, to the eventual formation of an eye with a single lens focusing images onto a retina. In humans and other vertebrates, the retina consists not only of photoreceptor cells but also of several types of neurons that begin to analyze the visual image. Through such gradual steps, very different kinds of eyes have evolved, from simple light-sensing organs to highly complex systems for vision.
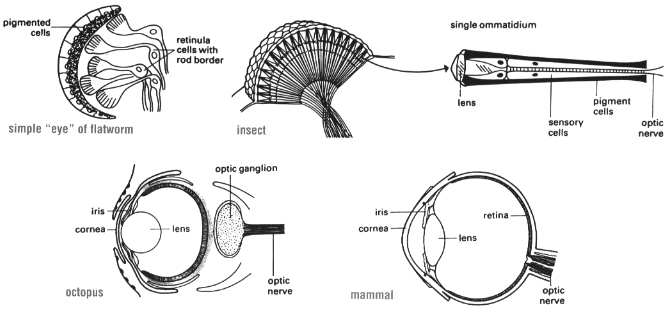
Eyes evolved over many millions of years from simple organs that can detect light.














