
2
Variola Virus and Other Orthopoxviruses
Viruses form a distinct group of infectious agents that are fundamentally different from bacteria and protozoa. The infectious particle, called the virion, requires the machinery of a host's living cells to reproduce. Viruses become active only after entering a host cell either by membrane fusion (enveloped viruses) or by a process that ''uncoats" the virus. The latter process causes the virus to shed some of its outer components so its inner core of genetic material has access to components of the host cell in order to be transcribed and translated. The accepted classification of viruses is based primarily on the morphology of the virion and the nature and structure of the viral nucleic acid. The primary taxonomic division consists of two classes based on nucleic acid content: DNA viruses and RNA viruses. Both categories are further subdivided into viruses that contain either double-stranded or single-stranded DNA or RNA.
The basic taxonomic group is called a family, designated by the "viridae" suffix. The family Poxviridae contains the largest of all viruses; the virions of the poxviruses are the only virus particles that can be seen with a light microscope. The virions have an ovoid or brick-like shape with dimensions of 400 by 200 nanometers and a linear genome of double-stranded DNA of sufficient length to encode approximately 200 proteins. There are two subfamilies: Chordopoxvirinae , which infect vertebrates, and Entomopoxvirinae, which infect arthropods. Replication in the cytoplasm of the host cell and the presence of viral enzymes for replication and expression of their genome are characteristic features of all poxviruses. The Chordipoxvirinae are divided into subgroups based on morphology, host range, and serological cross-reactivity.
General Attributes of Orthopoxviruses
The genus Orthopoxvirus is relatively more homogeneous than other members of the Chordopoxvirinae subfamily and includes 11 distinct but closely related species showing extensive serological cross-reactivity (see Table 2-1). Neutralization and cross-protection in laboratory animals form the original basis for defining the genus.
The external surface of the poxvirions is ridged in parallel rows that are sometimes arranged helically (see Figure 2-1). As noted, the viral particles contain approximately 200 proteins, about half structural and half nonstructural. Their internal structure is complex. Viewing thin negative-stained sections of virions in an electron microscope reveals that the outer surface is composed of lipid and protein that surrounds the core, which is biconcave (dumbbell-shaped) with two "lateral bodies" of unknown function. The core is composed of tightly compressed nucleoprotein. Poxviruses contain both specific and common proteins. The common proteins induce cross-reactive immunity and account for the ability to vaccinate against disease from another poxvirus of the same genus. There are at least 10 enzymes present in the particle that mediate gene expression. Of all the poxviruses, only those of the genus Orthopoxvirus produce a hemagglutinin antigen (HA).
TABLE 2-1 Species of the Genus Orthopoxvirus
|
Species |
Animals Infected |
Host Range |
Geographic Range |
|
Variola |
Human |
Narrow |
Formerly worldwide |
|
Vaccinia |
Human,a cow, pig, buffalo, rabbit, etc. |
Broad |
Worldwideb |
|
Cowpox |
Rodent,a cow, human, cat, etc. |
Broad |
Europe |
|
Monkeypox |
Squirrel,a monkey, ape, human |
Broad |
Western and central Africa |
|
Ectromelia |
Mouse, mole |
Narrow |
Europe |
|
Camelpox |
Camel |
Narrow |
Africa and Asia |
|
Taterapox |
Gerbil |
Narrow |
Western Africa |
|
Volepox |
Vole |
? |
United States |
|
Raccoonpox |
Raccoon |
? |
United States |
|
Skunkpox |
Skunk |
? |
United States |
|
Uasin Gishu |
Horse |
Medium |
Eastern Africa |
|
a Primary host. b Secondary to vaccination; no known natural host. |
|||
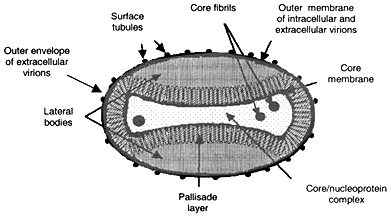
Figure 2-1
Stylized poxvirion from negative stain images [3].
Poxvirus Replication
Poxviruses can reproduce only within a host cell. The infection process produces many progeny that are replicas of the parent virus. The first step in the cycle of infection is attachment of the invading virion to the surface of the host cell. Usually the viral DNA genetic material cannot be replicated until it is released from the virion into the host cell.
Most of what is known about poxvirus intracellular growth has come from studies of vaccinia virus. There are two different infectious forms of vaccinia virus—Intracellular Mature Virus (IMV) and Extracellular Enveloped Virus (EEV)—whose virions differ in their role in the virus life cycle, their interaction with the immune system, and the way they bind to and enter cells. However, the question of the relative importance of the IMV and EEV forms for variola virus remains unanswered.
For poxviruses, the synthesis of messenger RNA begins before the genome is uncoated, and is moderated by RNA polymerase and other enzymes packaged within the infectious particle. The early messenger RNA is translated into proteins that facilitate the uncoating and replication of the genome and allow transcription of a second class of intermediate genes. The intermediate messenger RNA is translated into factors that allow transcription of the late class of genes. The late messenger RNA is translated into the structural and enzyme components of the virion. The newly replicated progeny genomes are incorporated into the virions being assembled. In culture, the last step of the growth cycle is the exit of progeny virions by eventual lysis or less frequently by exocytosis, in which virions are enveloped by virus-altered cell membranes of the host cell. The progeny virions can then spread the infection to neighboring cells, to other
sites, or to other individuals [1, 10]. Viremia in vivo is largely cell-associated, and mechanisms of spread are not fully understood.
Within several hours of infection, so-called "toxic," or cytopathic, changes occur in infected cells. This process involves general shutdown of the synthesis of host DNA, RNA, and protein, as well as changes in the cell architecture. These changes occur as the virus takes over the metabolic machinery of the cell for its own purposes.
Properties of Specific Orthopoxviruses
Poxviruses contain large double-stranded genomes and differ from most other DNA viruses in that they replicate in the cytoplasm rather than in the nucleus of susceptible cells. Selected properties of five orthopoxvirus species are described here because of the relevance of these species to the assessment of future scientific needs for live variola virus.*
Variola Virus. Variola is a human-specific virus. Generally it can be readily distinguished from other orthopoxviruses capable of infecting man (vaccinia, cowpox, monkeypox) by the characteristic small white pocks produced on the chorioallantoic membrane of developing 12-to 15-day-old chick embryos and the ceiling temperature of growth.
How, where, or when variola virus originated is not known. The basic question regarding the origin of a human-specific virus is how long ago, in terms of biological evolution, the viral species in question developed the capacity to be maintained indefinitely through human-to-human spread. Indefinite maintenance of a virus in populations of various sizes depends on three factors: (1) certain characteristics of the virus, notably its capacity to undergo antigenic change; (2) characteristics of the pathogenesis of the infection, especially the quality of the immune response and whether persistent infection or recurrence of infectivity occurs; and (3) characteristics of the population biology of the host, notably the rate of accession of new susceptible subjects. It has been estimated that a population of about 200,000 susceptible individuals would be necessary to support a sustained infection with humans as the sole host, and that this circumstance could have occurred only after the introduction of irrigated agriculture, about 10,000 years ago, had engendered the first great population explosion.
Two possibilities could explain the existence of a human-specific virus that has been recognized for 2,000 to 3,000 years. First, humans could have acquired the virus from some animal host in which the virus could be maintained in
larger numbers and had a much shorter generation time than in humans. Second, humans (or perhaps protohumans) may long have been the host of an ancestral "variola virus" that produced a different sort of disease that could persist in small groups of hunter-gatherers. The first of these possibilities 'appears probable. While monkeypox is a likely candidate since it causes a disease in humans that is very similar to smallpox, the variola virus genome exhibits greater similarity to other orthopoxviruses.
From the time it was first recognized until the end of the 19th century, smallpox was regarded as a uniformly severe disease with case-fatality rates in unvaccinated individuals of up to 40 percent. As noted in Chapter 1, in about 1900 a milder form of smallpox was identified with case-fatality rates of 1 percent or less. This milder form of smallpox is called variola minor, while the term variola major refers to the classical, more serious form. A more detailed description of the pathogenesis of and immune response to variola virus infections is presented in Chapter 3.
Vaccinia Virus. Vaccinia virus has been propagated by humans for use as a smallpox vaccine for the past 200 years. There are no known natural hosts of vaccinia virus, and its origin is obscure. All present strains of vaccinia virus are genetically related, although they exhibit different biological properties in the laboratory. Most strains have a wide host range in experimental animals, and all exhibit rapid growth on the chorioallantoic membrane cultures. Since the early 1980s, strains of vaccinia have been engineered to express genetic information for other viral and bacterial antigens or immunomodulatory proteins. Vaccination against smallpox is most effective by infection of the skin with vaccinia virus, followed by extension of replication to the lymph nodes and spleen, which elicits cell-mediated and humoral immune responses that provide protection against infection with variola virus.
The clinical events following primary inoculation of the skin with vaccinia virus are more rapid than the course of a natural smallpox infection, and are localized rather than generalized. Inoculation produces a papule at the vaccination site on the third day after vaccination. This papule becomes vesicular within 2 or 3 days. As with smallpox, the vesicle soon becomes pustular, mostly as a result of the entry of polymorphonuclear cells, the influx of which is induced by the viral infection. The surrounding skin becomes much more tender than in smallpox, and the lesions reach maximum size between 8 and 12 days after infection. At this time the lymph nodes are more or less enlarged and tender, and the subject sometimes experiences mild fever and malaise. The pustule dries from the center outward to become a scab that falls off about 3 weeks after vaccination to leave a typical pitted scar.
Monkeypox Virus. Human monkeypox is a systemic disease with a generalized pustular rash that is usually clinically indistinguishable from that of
smallpox. Monkeypox virus was discovered in 1958 when it was isolated from the lesions of captive monkeys at the State Serum Institute in Copenhagen. Monkeypox virus has a broad host range that includes most common laboratory animals. Monkeypox antibodies have been found in various wild animals, especially squirrels in Zaire.
The first case of human monkeypox was identified in Zaire in 1970, followed by four cases in Liberia and one in Sierra Leone. Clinically, human monkeypox closely resembles ordinary smallpox. The most obvious clinical difference is a pronounced lymph-node enlargement with monkeypox, which typically occurs in the neck and groin, but can be more generalized. Swollen lymph glands and fever precede the rash in most cases.
Prospective studies in Zaire between 1981 and 1986 have indicated that three-fourths of human monkeypox infections are attributable to direct contact with animals; the remainder can be traced to contact with infected persons. Whereas the person-to-person transmission rate to unvaccinated individuals in close contact ranges up to 70 percent for smallpox, it is about 8 percent for monkeypox. Serial transmission of monkeypox involving more than two or three people is rare. While monkeypox probably has existed in the tropical rain forests of Africa for a long time, it apparently has never established a continuous person-to-person infection in a human population. Preliminary results from sequencing of DNA fragments of monkeypox virus isolates obtained at various times since 1970 suggest that the virus has changed very little during this period. At present, there is no clear evidence that the rate of human-to-human transmission of monkeypox is likely to increase.*
Cowpox Virus. Cowpox is of interest primarily because of its role in the discovery of vaccination. The occurrence of a sporadic pox disease in cows that is transmissible to humans had been known for centuries when Edward Jenner, an English country physician, brought it to public attention (see the discussion of"Immunity Against Smallpox" in Chapter 3). The virus can infect a variety of animals, and is probably maintained over time in rodents. Its genome is the largest of all the orthopoxviruses, and deletion mutations occur commonly, producing progeny with smaller genomes. None of the strains of cowpox virus that have been examined looks at all like that of variola virus.
Mousepox (Infectious Ectromelia) Virus. This virus, discovered in 1930, is a natural pathogen of mice, causing serious disease with a rash in outbred and several inbred strains. It has been used to study the molecular basis of the virulence of orthopoxviruses since it has the advantage that the mouse, by far the best experimental animal for such studies, is its natural host [11]. Mousepox,








